
The Wobble Hypothesis: Importance and Examples
The “wobble hypothesis” refers to a concept in molecular biology that explains the degeneracy of codons.
So, what are codons? Codons are sets of three nucleotides in mRNA (messenger RNA) that correspond to specific amino acids. 64 possible codons codes for the 20 standard amino acids used in protein synthesis.
Since there are only 20 amino acids and 64 possible codons, multiple codons may code for a single amino acid during protein synthesis. In molecular biology, this redundancy or multiplicity of codons is termed degeneracy.
The wobble hypothesis or wobble theory, proposed by Francis Crick in 1966, suggests that the third base of a codon can sometimes be flexible or “wobble.” The wobbling or flexibility allows for non-standard base pairing between the mRNA codon and the tRNA (t RNA) anticodon during translation.
The first two nucleotides of the codon typically adhere to strict base-pairing rules. Still, the third position may tolerate mismatches, allowing for variations such as G-U (guanine-uracil) pairing or other non-standard interactions.
This hypothesis helps explain how a relatively limited number of tRNA molecules can recognize and bind to multiple codons for the same amino acid, facilitating efficient and accurate protein synthesis. Experimental evidence has supported the wobble hypothesis, a fundamental concept in understanding the genetic code and translation machinery.\

Table of Contents
Crick’s wobble hypothesis states that the base at the 5′ end of the anticodon does not confine spatially as the other two bases, which allows the development of hydrogen bonds with other bases present at the 3′ end of a codon. The wobble hypothesis outlines several key points:
- Degeneracy of the Genetic Code: The genetic code degenerates, meaning multiple codons can code for the same amino acid. For example, six codons, UUA, UUG, CUU, CUC, CUA, and CUG, code the amino acid leucine.
- Flexibility in Codon-Anticodon Interactions: The wobble hypothesis suggests that the base pairing between the mRNA codon’s third nucleotide and the tRNA anticodon’s corresponding nucleotide is flexible. Instead, it allows for some flexibility or “wobble” in the pairing.
- Non-Standard Base Pairing: The third position of the codon-anticodon interaction can tolerate non-standard base pairs, such as G-U (guanine-uracil) pairing or other non-Watson-Crick interactions. For example, a tRNA with the anticodon 3′-CCU-5′ can recognize the codons CGU, CGC, and CGA, where the third position allows for wobble pairing.
Importance of Wobble Hypothesis
The wobble hypothesis is essential in molecular biology for several reasons:
- Efficient Translation : The wobble hypothesis explains how fewer tRNA molecules can recognize multiple codons coding for the same amino acid. This reduces the number of tRNA species required for protein synthesis, streamlining the translation process and making it more efficient.
- Error Reduction : By allowing for flexibility in base pairing at the third position of the codon-anticodon interaction, the wobble hypothesis helps reduce the impact of errors or mutations in the genetic code. Even if a mutation occurs in the third position of a codon, it may not necessarily result in a change in the protein’s amino acid sequence, thereby minimizing errors in protein synthesis.
- Evolutionary Conservation : The wobble hypothesis is evolutionarily conserved across species, indicating its fundamental importance in translation. This conservation suggests that the wobble base pairing mechanism provides an evolutionary advantage by allowing for greater adaptability and efficiency in protein synthesis.
- Understanding Genetic Code Variability : The wobble hypothesis helps us understand the variability in the genetic code, where multiple codons can code for the same amino acid. This variability provides flexibility and redundancy in the genetic code. This allows for robustness and adaptability in the face of genetic mutations and environmental changes.
- Biotechnological Applications : Understanding the wobble hypothesis is crucial in biotechnology and genetic engineering applications. For example, it informs the design of synthetic genes and optimization of codon usage to enhance protein expression in heterologous expression systems.
Overall, the wobble hypothesis plays a fundamental role in understanding protein synthesis and the genetic code, with implications for various aspects of molecular biology, genetics, and biotechnology.
Examples of Wobble Hypothesis
Here are some examples of the wobble hypothesis in action:
- Arginine: The amino acid arginine is coded by six different codons: CGU, CGC, CGA, CGG, AGA, and AGG. However, no six different tRNA molecules correspond to each of these codons. Instead, one tRNA molecule with the anticodon 3′-CCU-5′ can recognize the codons CGU, CGC, and CGA (where the third nucleotide is flexible), thanks to wobble base pairing.
- Leucine: Leucine is another example of wobble base pairing. The codons UUA, UUG, CUU, CUC, CUA, and CUG are all codes for leucine. However, the tRNA molecule with the anticodon 3′-AAG-5′ can recognize UUA and UUG codons due to wobble base pairing at the third position.
- Serine: Serine is encoded by six codons: UCU, UCC, UCA, UCG, AGU, and AGC. Due to wobble base pairing, the tRNA molecule with the anticodon 3′-AGU-5′ can recognize both AGU and AGC codons.
- Isoleucine: The codons AUU, AUC, and AUA all code for isoleucine. The tRNA molecule with the anticodon 3′-IAU-5′ (where “I” represents inosine, a modified nucleotide capable of wobble base pairing) can recognize all three codons through wobble interactions.
These examples illustrate how the wobble hypothesis allows for flexibility in the genetic code, enabling fewer tRNA molecules to recognize multiple codons and facilitating efficient protein synthesis.
Limitation of Wobble Hypothesis
While the wobble hypothesis provides a valuable framework for understanding how the genetic code is flexible and the efficiency of translation, it also has some limitations and considerations:
- Context-dependence : The wobble hypothesis primarily applies to the standard codon-anticodon interactions during translation. However, non-standard base pairing beyond the wobble hypothesis may occur in certain contexts or under specific conditions. For example, modified nucleotides in tRNA or mRNA can influence base pairing interactions in ways that go beyond traditional wobble pairing rules.
- Accuracy and Specificity : While wobble base pairing can contribute to the recognition of multiple codons by a single tRNA molecule, it may also lead to potential errors during translation. The flexibility in the third position of the codon-anticodon interaction could allow non-standard base pairs to form. This can potentially lead to misinterpretation of the genetic code and errors in protein synthesis.
- Influence of Structural Constraints : The wobble hypothesis primarily focuses on the base pairing interactions between codons and anticodons. However, other factors such as tRNA structure, modifications, and interactions with the ribosome also influence the accuracy and efficiency of translation. These factors may impose additional constraints or considerations beyond the wobble hypothesis.
- Evolutionary Variability : While the wobble hypothesis explains a general trend in codon-anticodon recognition, there can be variations in wobble base pairing preferences across species or even within different tissues or cellular conditions. Evolutionary pressures, genetic variations, and differences in tRNA modifications can influence the extent and specificity of wobble interactions.
- Complexity of Codon Usage : The relationship between codon usage bias, tRNA abundance, and wobble interactions is complex and can vary between organisms and genes. While wobble base pairing contributes to codon redundancy and efficient translation, other factors such as codon optimality, mRNA secondary structure, and ribosome kinetics influence translation efficiency and protein expression levels.
- Crick F. H. (1966). Codon–anticodon pairing: the wobble hypothesis. Journal of molecular biology , 19 (2), 548–555. https://doi.org/10.1016/s0022-2836(66)80022-0
- Mangang, S. U., & Lyngdoh, R. H. (2001). Wobble base-pairing in codon-anticodon interactions: a theoretical modelling study. Indian journal of biochemistry & biophysics , 38 (1-2), 115–119.
- Verma, P. S., & Agarwal, V. K. (2019). Cell Biology, genetics, Molecular Biology, evolution and ecology (25th ed.). S. Chand and Company Limited.
Ashma Shrestha
Hello, I am Ashma Shrestha. I had recently completed my Masters degree in Medical Microbiology. Passionate about writing and blogging. Key interest in virology and molecular biology.
We love to get your feedback. Share your queries or comments Cancel reply
This site uses Akismet to reduce spam. Learn how your comment data is processed .
Recent Posts
Urea Cycle: Steps, End Products, and Functions
Amino nitrogen, a key component in the synthesis of amino acids or new nitrogenous products, can be toxic to the human body if not utilized to create new compounds. To prevent this, ureotelic...
Gluconeogenesis: Enzymes Involved, Steps, and Functions
During fasting, vigorous exercise, and hypoglycemic conditions, the body requires high glucose. Gluconeogenesis converts non-carbohydrate molecules like glycerol, pyruvate, lactate, glucogenic amino...

- BiologyDiscussion.com
- Follow Us On:
- Google Plus
- Publish Now
Wobble Hypothesis (With Diagram) | Genetics

ADVERTISEMENTS:
In this article we will discuss about the concept of wobble hypothesis.
Crick (1966) proposed the ‘wobble hypothesis’ to explain the degeneracy of the genetic code. Except for tryptophan and methionine, more than one codons direct the synthesis of one amino acid. There are 61 codons that synthesise amino acids, therefore, there must be 61 tRNAs each having different anticodons. But the total number of tRNAs is less than 61.
This may be explained that the anticodons of some tRNA read more than one codon. In addition, identity of the third codon seems to be unimportant. For example CGU, CGC, CGA and CGG all code for arginine. It appears that CG specifies arginine and the third letter is not important. Conventionally, the codons are written from 5′ end to 3′ end.
Therefore, the first and second bases specify amino acids in some cases. According to the Wobble hypothesis, only the first and second bases of the triple codon on 5′ → ‘3 mRNA pair with the bases of the anticodon of tRNA i.e A with U, or G with C.
The pairing of the third base varies according to the base at this position, for example G may pair with U. The conventional pairing (A = U, G = C) is known as Watson-Crick pairing (Fig. 7.1) and the second abnormal pairing is called wobble pairing.
This was observed from the discovery that the anticodon of yeast alanine-tRNA contains the nucleoside inosine (a deamination product of adenosine) in the first position (5′ → 3′) that paired with the third base of the codon (5′ → 3′). Inosine was also found at the first position in other tRNAs e.g. isoleucine and serine.
The purine, inosine, is a wobble nucleotide and is similar to guanine which normally pairs with A, U and C. For example a glycine-tRNA with anticodon 5′-ICC-3′ will pair with glycine codons GGU, GGC, GGA and GGG (Fig 7.2). Similarly, a seryl-tRNA with anticodon 5′-IGA-3′ pairs with serine codons UCC, UCU and UCA (5-3′). The U at the wobble position will be able to pair with an adenine or a guanine.

According to Wobble hypothesis, allowed base pairings are given in Table 7.5:

Due to the Wobble base pairing one tRNA becomes able to recognise more than one codons for an individual amino acid. By direct sequence of several tRNA molecules, the wobble hypothesis is confirmed which explains the pattern of redundancy in genetic code in some anticodons (e.g. the anticodons containing U, I and G in the first position in 5’→ 3′ direction)

The seryl-tRNA anticodon (UCG) 5′-GCU-3′ base pairs with two serine codons, 5′-AGC-3′ and 5′-AGU-3′. Generally, Watson-Crick pairing occurs between AGC and GCU. However, in AGU and GCU pairing, hydrogen bonds are formed between G and U. Such abnormal pairing called ‘Wobble pairing’ is given in Table 7.5.
Three types of wobble pairings have been proposed:
(i) U in the wobble position of the tRNA anticodon pairs with A or G of codon,
(ii) G pairs with U or C, and
(iii) 1 pairs with A, U or C.
Related Articles:
- Short Notes on Anticodons | Genetics
- Genetic Code: Degeneracy and Universality | Protein
Microbiology , Genetics , Wobble Hypothesis , Concept of Wobble Hypothesis
- Anybody can ask a question
- Anybody can answer
- The best answers are voted up and rise to the top
Forum Categories
- Animal Kingdom
- Biodiversity
- Biological Classification
- Biology An Introduction 11
- Biology An Introduction
- Biology in Human Welfare 175
- Biomolecules
- Biotechnology 43
- Body Fluids and Circulation
- Breathing and Exchange of Gases
- Cell- Structure and Function
- Chemical Coordination
- Digestion and Absorption
- Diversity in the Living World 125
- Environmental Issues
- Excretory System
- Flowering Plants
- Food Production
- Genetics and Evolution 110
- Human Health and Diseases
- Human Physiology 242
- Human Reproduction
- Immune System
- Living World
- Locomotion and Movement
- Microbes in Human Welfare
- Mineral Nutrition
- Molecualr Basis of Inheritance
- Neural Coordination
- Organisms and Population
- Photosynthesis
- Plant Growth and Development
- Plant Kingdom
- Plant Physiology 261
- Principles and Processes
- Principles of Inheritance and Variation
- Reproduction 245
- Reproduction in Animals
- Reproduction in Flowering Plants
- Reproduction in Organisms
- Reproductive Health
- Respiration
- Structural Organisation in Animals
- Transport in Plants
- Trending 14
Privacy Overview

- Collections
Wobble Hypothesis

Wobble Base Pair
- A wobble base pair is type of non-canonical base pairing that occurs between two nucleotides in RNA molecules, the codon and the anticodon of mRNA and tRNA, respectively that does not follow Watson-Crick base pair rules.
- The four main wobble base pairs are guanine-uracil ( G-U ), hypoxanthine-uracil ( I-U ), hypoxanthine-adenine ( I-A ), and hypoxanthine-cytosine ( I-C )
- Wobble base pairs are fundamental in RNA secondary structure and are critical for the proper translation of the genetic code
- The term “wobble” refers to the flexibility or deviation from the standard Watson-Crick base pairing rules at the third position of the codon, which allows a single tRNA to recognize more than one codon for the same amino acid.
- This phenomenon explains the degeneracy of the genetic code and reduces the number of tRNA molecules required for protein synthesis.
The Wobble Hypothesis
Genetic Code Study Notes:
- There are 64 possible codons in the genetic code, each consisting of a 3-nucleotide sequence. Translation requires tRNA molecules, each with an anticodon that complements a specific mRNA codon. Canonical Watson-Crick base pairing is used for stable tRNA-mRNA binding during translation.
- In the standard genetic code, 3 mRNA codons (UAA, UAG, UGA) act as stop codons, terminating translation. This leaves 61 mRNA codons that require tRNA molecules, suggesting a need for 61 types of tRNA.
- Due to the limited number of tRNA species in organisms (usually fewer than 45), some tRNA types can pair with multiple synonymous codons.
- Francis Crick proposed the Wobble Hypothesis in 1966, suggesting that the 5′ base on the anticodon has non-standard base pairing due to spatial flexibility. The “wobble” at the third codon position allows for small conformational adjustments, influencing the overall pairing geometry of tRNA anticodons.
- Crick suggested that the first two bases of the codon form strong and specific Watson-Crick base pairs with the second and third bases of the anticodon, while the third base of the codon can form weaker and less specific base pairs with the first base of the anticodon.
- The first two bases of each codon are primary determinants of specificity. The third base pairing is not very stable and wobbles. For example, CUU, CUG, CUC, CUA codons, which differ only at the third base represent the same amino acid leucine. The first two bases of the codon form strong base pairs with the corresponding bases of the anticodon but the third base forms weak hydrogen bond.
- This allows some tRNA molecules to bind to more than one codon, as long as they differ only at the third position. For example, a tRNA with the anticodon 5′-GmAA-3′ can recognize both UUC and UUU codons for phenylalanine.
The Wobble Rules
Crick also proposed a set of rules that govern the possible wobble base pairs, based on the geometry and hydrogen bonding patterns of the nucleotides involved. The rules are as follows:
- G can pair with U or C (in addition to the canonical C)
- U can pair with A or G (in addition to the canonical A)
- I (inosine, a deaminated form of A) can pair with A, U, or C
- A can pair only with U (the canonical pair)
- C can pair only with G (the canonical pair)
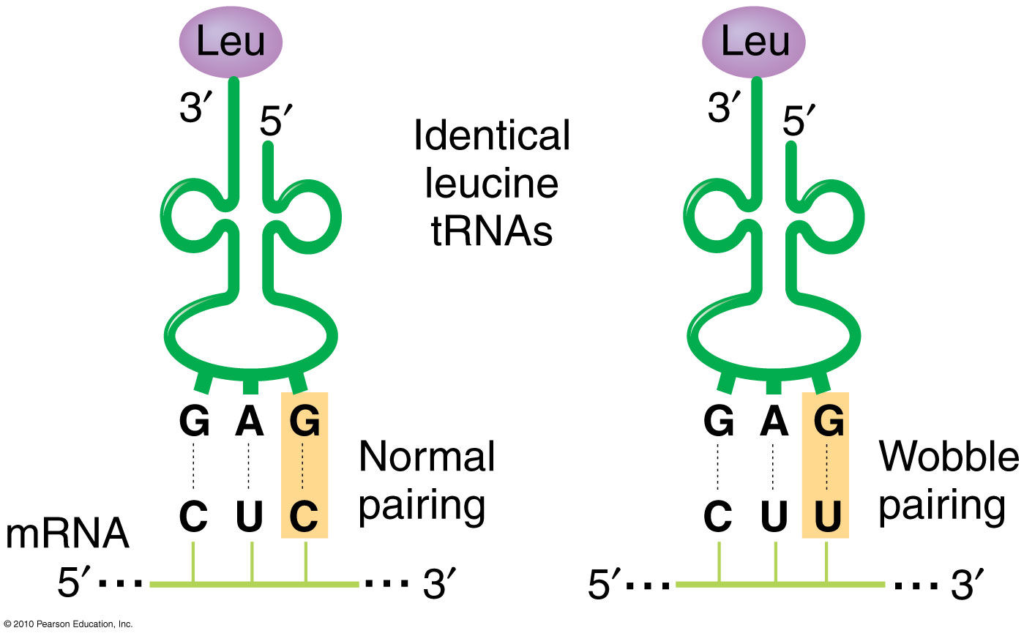
These rules imply that some codons for the same amino acid are more versatile than others in terms of wobble pairing. For instance, codons ending with A or C can be recognized by only one specific tRNA, while codons ending with U or G can be recognized by one or two tRNAs, depending on whether the first base of the anticodon is U, G, or I.
The Significance of Wobble Base Pairing
Wobble base pairing has several advantages for the cell:
- It reduces the number of tRNA genes and tRNA molecules needed to translate all 61 sense codons, which saves genetic space and energy.
- It allows for some mutations or errors at the third position of the codon without affecting the protein sequence, which increases the genetic robustness and diversity.
- It enables some tRNA molecules to act as suppressors of nonsense mutations by recognizing stop codons and inserting amino acids instead, which may rescue some defective proteins.
Wobble base pairing is a common feature of RNA secondary structure and is essential for the accurate and efficient translation of the genetic code.
Sign Up For Daily Newsletter
Be keep up get the latest articles delivered straight to your inbox..
I have read and agree to the terms & conditions
Leave a review Cancel reply
Your email address will not be published. Required fields are marked *
Your comment *
Your name *
Your Email *
Captcha Plus loading... In order to pass the CAPTCHA please enable JavaScript.
Biotechtutorials
Share knowledge..
- Quick Links
Username or Email Address
Remember Me
Update image
- Wobble hypothesis

Support our developers

More in this section
- The Genetic Code
- Properties of genetic code
- Chain initiation and chain termination codons
- Synonym codons and degeneracy
- Mutations and the genetic code
- New genetic codes in mitochondria and ciliate protozoa
- Suppressor mutations, base substitutions and suppressor tRNAs
- Second genetic code, and second half of the genetic code
- Recoding of the genetic code
© 2012 - 2024 Biocyclopedia
Disclaimer Privacy Policy Feedback
Login to your account
Login / signup with.


- __Crossword/Puzzle
- __Resources
- Difference Between
- Biology MCQ
- _Chemistry for Biologists
What is Wobble Hypothesis?
- The multiple codes for a given amino acid( degeneracy )
- Possible suppression of point mutations in the third base of the codon
- More easily removed deacylated tRNA during protein synthesis.
- Fewer tRNA molecules than expected.
Our website uses cookies to improve your experience. Learn more
Contact form

Microbiology Notes
Genetic Code – Definition, Characteristics, Wobble Hypothesis
Table of Contents
What is a Genetic Code?
The genetic code is a set of rules that living cells use to decipher the information encoded in genetic material (DNA or mRNA sequences). The ribosomes are responsible for carrying out the translation process. Using tRNA (transfer RNA) molecules to carry amino acids and to read the mRNA three nucleotides at a time, they link the amino acids in an mRNA-specified (messenger RNA) order.
- As DNA is a genetic substance, it transmits genetic information from one cell to the next and from one generation to the next.
- At this point, it will be attempted to determine how genetic information is stored within the DNA molecule. On the DNA molecule, are they written in an articulated or encoded language? In the language of codes, what is the genetic code’s nature?
- A DNA molecule contains three types of moieties: phosphoric acid, deoxyribose sugar, and nitrogen bases.
- The genetic information may be encoded in any of the three DNA molecules. However, because the poly-sugarphosphate backbone is always the same, it is doubtful that these DNA molecules convey genetic information.
- However, the nitrogen bases vary from one DNA segment to the next, therefore the information may depend on their sequences.
- In fact, the sequences of nitrogen bases in a specific section of DNA are similar to the linear sequence of amino acids in a protein molecule.
- An investigation of mutations of the head protein of bacteriophage T4 and the A protein of tryptophan synthetase from Escherichia coli provided the initial evidence for the colinearity between DNA nitrogen base sequence and amino acid sequence in protein molecules.
- Colinearity between protein molecules and DNA polynucleotides provides evidence that the arrangement of four nitrogen bases (e.g., A, T, C, and G) in DNA polynucleotide chains dictates the sequence of amino acids in protein molecules.
- These four DNA bases can therefore be viewed as the four alphabets of the DNA molecule. Therefore, all genetic information should be encoded using these four DNA alphabets.
- The question that now emerges is whether genetic information is written in articulated or coded language. If genetic information could have been communicated in an articulated language, the DNA molecule would have required multiple alphabets, a complicated grammar system, and adequate space.
- All of these could be practically difficult and also problematic for the DNA. Therefore, it was reasonable for molecular biologists to assume that genetic information resided in the DNA molecule as a specific language of code words that utilised the four nitrogen bases of DNA as their symbols. Any encoded message is referred to as a cryptogram.

Basis of Cryptoanalysis
- How information written in a four-letter language (four nucleotides or nitrogen bases of DNA) may be transformed into a twenty-letter language is the fundamental challenge of such a genetic code (twenty amino acids of proteins).
- A code word or codon is the set of nucleotides that specifies one amino acid. By genetic code, one refers to the collection of sequences of bases (codons) that correspond to each amino acid and translation signals.
- Regarding the possible size of a codon, we can consider George Gamov’s (1954) traditional yet rational explanation.
- The simplest conceivable code is a singlet code (a code of a single letter) that specifies a single nucleotide amino acid.
- A doublet code (consisting of two letters) is similarly insufficient, as it can only define sixteen (4×4) amino acids, but a triplet code (consisting of three letters) can specify sixty-four (4x4x4) amino acids.
- Therefore, it is probable that 64 triplet codes exist for 20 amino acids. The conceivable singlet, doublet, and triplet codes, which are conventionally described in terms of “mRNA language” [mRNA is a complementary molecule that copies the genetic information (cryptogram of DNA) during its transcription] are depicted in Table.
- In 1961, Crick and his colleagues present the first experimental evidence supporting the hypothesis of triplet coding.
- During their experiment, when they inserted or deleted single or double base pairs in a specific region of the DNA of E.coli T4 bacteriophages, they discovered that these bacteriophages ceased to execute their regular tasks.
- Nevertheless, bacteriophages with the addition or deletion of three base pairs in the DNA molecule had normal functionality.
- In this experiment, the addition of one or two nucleotides caused the message to be read incorrectly, however the addition of a third nucleotide resulted in the message being read correctly again.

Codon Assignment (Cracking the Code or Deciphering the Code)
The genetic code has been broken or deciphered using the following methods:
A. Theoretical Approach
- George Gamow, a physicist, proposed the diamond code (1954) and the triangle code (1955), as well as a comprehensive theoretical framework for the various aspects of the genetic code.
- A triplet codon that corresponds to a single polypeptide chain amino acid.
- Direct template translation by linking codons with amino acids.
- The code is translated in an overlapping fashion.
- Degeneration of the code, or the coding of an amino acid by more than one codon.
- The colinearity of nucleic acid and the produced main protein.
- Universality of the code, i.e., the code being fundamentally identical throughout organisms.
- Molecular biologists have refuted a number of these statements by Gamow. Brenner (1957) demonstrated that the overlapping triplet code is impossible, and further research has demonstrated that the code is non-overlapping.
- Crick’s adopter hypothesis similarly contested Gamow’s assumption of a direct template relationship between nucleic acid and polypeptide chain.
- Adaptor molecules, according to this concept, intervene between nucleic acid and amino acids during translation.
- In actuality, it is now understood that tRNA molecules serve as adaptors between the codons of mRNA and the amino acids of the resultant polypeptide chain.
B. The in vitro codon Assignment
1. discovery and use of polynucleotide phosphorylase enzyme.
Marianne Grunberg Manago and Severo Ochoa identified an enzyme from bacteria (e.g., Azobacter vinelandii or Micrococcus lysodeikticus) that catalyses RNA degradation in bacterial cells. The name of this enzyme is polynucleotide phosphorylase. Outside of the cell (in vitro), with high amounts of ribonucleotides, Manago and Ochoa discovered that the reaction could be driven in reverse and an RNA molecule could be produced (see Burns and Bottino, 1989). The random incorporation of nucleotides into the molecule is independent of a DNA template. Thus, in 1955, Manago and Ochoa made possible the artificial synthesis of polynucleotides (=mRNA) comprising only a single type of nucleotides (U, A, C, or G, respectively, repeated several times).

Consequently, the action of polynucleotide phosphorylase can be depicted as follows:

The polynucleotide phosphorylase enzyme differs from RNA polymerase used to transcribe mRNA and DNA polymerase used to transcribe mRNA from DNA in the following ways: I it does not require a template or primer; (ii) the activated substrates are ribonucleoside diphosphates (e.g., UDP, ADP, CDP, and GDP) and not triphosphates; and (iii (PPi). The introduction of synthetic (or artificial) polynucleotides and trinucleotides made the deciphering of the genetic code possible.
Use of polymers containing a single type of nucleotide (called homopolymers), mixed polymers (copolymers) containing multiple types of nucleotides (heteropolymers) in random or defined sequences, and trinucleotides (or “minimessengers”) in ribosome-binding or filter-binding are among the various techniques employed.
2. Codon assignment with unknown sequence
(i) codon assignment by homopolymer..
- Marshall Nirenberg and Heinrich Matthaei (1961) supplied the first indication to codon assignment when they utilised an in vitro technique for the creation of a polypeptide utilising an artificially produced mRNA molecule containing only one type of nucleotide (i.e., homopolymer).
- Before doing the actual tests, they evaluated the capacity of a cell-free protein synthesis system to integrate radioactive amino acids into newly produced proteins.
- Their E.coli cell-free extracts comprised ribosomes, tRNAs, aminoacyl-tRNA synthetase enzymes, DNA, and messenger RNA.
- This extract’s DNA was eliminated by the deoxyribonuclease enzyme, so destroying the template for the synthesis of new mRNA.
- When twenty amino acids together with ATP, GTP, K+, and MG2+ were introduced to this mixture, they were integrated into proteins.
- As long as mRNA was present in the cell-free suspension, incorporation persisted. It also continued in the presence of synthetic polynucleotides (mRNAs) that might be synthesised using the polynucleotide phosphorylase enzyme.
- Nirenberg and Matthaei made the first successful application of this approach when they created a chain of uracil molecules (poly U) as their synthetic mRNA (homopolymer).
- A message consisting of a single base could not contain ambiguity, hence Poly (U) looked to be the best option. It binds well to ribosomes and, as it turned out, the resultant protein was insoluble and simple to isolate.
- When poly (U) was supplied as the message to the cell-free system containing all the amino acids, polyphenylalanine was picked solely from the mixture for incorporation into the polypeptide.
- This amino acid was phenylalanine, hence it was deduced that a sequence of UUU encoded for phenylalanine. Other homogeneous nucleotide chains (Poly A, Poly C, and Poly G) were inert for incorporation of phenylalanine. The phenlalanine mRNA code was consequently determined to be UUU.
- AAA is derived to be the equivalent DNA code word for phenylalanine. Thus, UUU was the first code word to be decrypted. In the laboratories of Nirenberg and Ochoa, this finding was developed.
- Using synthetic poly (A) and poly (C) chains, the experiment was repeated, yielding polylysine and polyproline, respectively.
- Thus, AAA was determined to be the code for lysine and CCC was determined to be the code for proline. A poly (G) message was discovered to be nonfunctional in vitro due to its secondary structure, which prevented it from attaching to ribosomes. Thus, three of the sixty-four codons were simply explained.
(ii) Codon assignment by heteropolymers (Copolymers with random sequences)
- Using synthetic messenger RNAs containing two different types of nucleotides, the genetic code was elucidated further.
- This approach was utilised in the laboratories of Ochoa and Nirenberg to deduce the codon composition for the 20 amino acids.
- The bases in the synthetic messengers were chosen at random (called random copolymers). In a random copolymer composed of U and A nucleotides, for instance, eight triplets are feasible, including UUU, UUA, UAA, UAU, AAA, AAU, AUU, and AUA.
- Theoretically, these eight codons may code for eight amino acids. However, actual experiments produced only six amino acids: phenylalanine, leucine, tyrosine, lysine, asparagine, and isoleucine.
- It was feasible to derive the composition of the code for different amino acids by altering the relative proportions of U and A in the random copolymer and determining the fraction of the different amino acids in the proteins generated.
3. Assignment of codons with known sequences.
- I The application of trinucleotides or minimessengers in filter binding (Ribosome-binding technique). Nirenberg and Leder’s (1964) ribosome binding technique takes use of the observation that aminoacyl-tRNA molecules attach selectively to the ribosomemRNA complex.
- The connection of a trinucleotide or minimessenger with the ribosome is necessary for aminoacyltRNA binding to occur.
- When a mixture of such small mRNA molecules-ribosomes and amino acid-tRNA complexes is incubated for a brief period and then filtered over a nitrocellulose membrane, the mRNA-ribosome-tRNA-amino acid complex is kept and the remainder of the mixture is discarded.
- Using a series of 20 different amino acid mixtures, each containing one radioactive amino acid, it is possible to determine the amino acid corresponding to each triplet by analysing the radioactivity absorbed by the membrane; for instance, the triplet GCC and GUU retain only alanyl-tRNA and valyl-tRNA, respectively.
- In this manner, all 64 potential triplets have been synthesised and evaluated. 45 of them have produced conclusive results. Later on, with the use of lengthier synthetic messages, 61 of the 64 potential codons have been deciphered.

C. The in vivo Codon Assignment
- Despite the fact that cell-free protein synthesis systems have played a significant role in the decipherment of the genetic code, they cannot tell us whether the deciphered genetic code is likewise utilised in the living systems of all organisms.
- Different molecular biologists use three techniques to determine if the same code is used in vivo: (a) amino acid replacement studies (e.g., tryptophan synthetase synthesis in E.coli and haemoglobin synthesis in man), (b) frameshift mutations (e.g., Terzaghi et al. 1966, on lysozyme enzyme of T4 bacteriophages), and (c) comparison of a DNA (e.g., comparison of amino acid sequence of the R17 bacteriophage coat protein with the nucleotide sequence of the R17 mRNA in the region of the molecule that dictates coat-protein synthesis by S. Cory et al., 1970).
- Thus, the previously mentioned in vitro and in vivo experiments allowed for the formulation of a code table for twenty amino acids.
Characteristics of Genetic Code
The genetic code has the following general properties :
1. The code is a triplet codon
- The nucleotides of messenger RNA (mRNA) are organised as a linear sequence of codons, with each codon consisting of three consecutive nitrogenous bases, i.e., the code is a triplet codon.
- Two types of point mutations, frameshift mutations and base substitution, provide support for the concept of triplet codon.
(i) Frameshift mutations
- Evidently, the genetic communication, once launched at a particular place, is decoded into a series of three-letter phrases within a specific time frame.
- As soon as one or more bases are removed or added, the structure would be disrupted. When such frameshift mutations were intercrossed, they produced wild-type normal genes in certain combinations.
- It was determined that one was a deletion and the other was an insertion, so that the disordered frame order caused by the mutation will be corrected by the other.
(ii) Base substitution
- If, at a specific location in an mRNA molecule, one base pair is replaced by another without deletion or insertion, the meaning of a codon containing the altered base will be altered.
- As a result, another amino acid will be inserted in place of a particular amino acid at a particular location in a polypeptide.
- Due to a substitution mutation in the gene for the tryptophan synthetase enzyme in E. coli, the glycine-coding GGA codon becomes the arginine-coding AGA.
- A missense codon is a codon that has been altered to specify a different amino acid. The discovery that a fragment of mRNA comprising 90 nucleotides corresponded to a polypeptide chain having 30 amino acids of a developing haemoglobin molecule provided more direct proof for the existence of a triplet code.
- Similarly, 1200 nucleotides of the “satellite” tobacco necrosis virus direct the creation of 372 amino acid-containing coat protein molecules.
2. The code is non-overlapping
- In the translation of mRNA molecules, codons are “read” sequentially and do not overlap.
- Therefore, a non-overlapping coding indicates that a nucleotide in an mRNA is not utilised for multiple codons.
- In practise, however, six bases code for no more than two amino acids. In the event of an overlapping code, for instance, a single change (of replacement type) in the base sequence will result in several amino acid substitutions in the associated protein.
- In insulin, tryptophan synthetase, TMV coat protein, alkaline phosphatase, haemoglobin, etc., a single base substitution leads in a single amino acid change. Since 1956, a large number of examples have accumulated in which a single base substitution results in a single amino acid change.
- Recently, however, it has been demonstrated that overlapping genes and codons are possible in bacteriophage φ × 174.
3. The code is commaless
- The genetic code is punctuation-free, thus no codons are reserved for punctuation.
- It means that when one amino acid is coded, the next three characters will automatically code the second amino acid and no letters will be wasted as punctuation marks.
4. The code is non-ambiguous
- A codon always codes for the same amino acid when it is non-ambiguous.
- In the situation of ambiguous code, the same codon may have many meanings; in other words, the same codon may code for two or more amino acids. As a general rule, a single codon should never code for two distinct amino acids.
- There are, however, documented exceptions to this rule: the codons AUG and GUG may both code for methionine as beginning or starting codons, despite the fact that GUG is intended for valine. Similarly, the GGA codon represents the amino acids glycine and glutamic acid.
5. The code has polarity
The direction in which the code is always read is 5’→3′. Thus, the codon possesses a polarity. Clearly, if the code is read in opposing directions, it would specify two distinct proteins, as the codon’s base sequence would be reversed:

6. The code is degenerate
- Multiple codons might define the same amino acid; this phenomenon is known as degeneracy of the code. Except for tryptophan and methionine, which each contain a single codon, the remaining 18 amino acids have several codons.
- Consequently, each of the nine amino acids phenylalanine, tyrosine, histidine, glutamine, asparagine, lysine, aspartic acid, glutamic acid, and cysteine has two codons. Isoleucine consists of three codons.
- Each of the five amino acids valine, proline, threonine, alanine, and glycine has four codons. Each of the three amino acids leucine, arginine, and serine has six codons.
- There are essentially two types of code degeneration: partial and total. Partial degeneracy occurs when the first two nucleotides of degenerate codons are identical, but the third (3′ base) nucleotide differs, e.g., CUU and CUC code for leucine.
- Complete degeneracy happens when any of the four bases can code for the same amino acid in the third position (e.g., UCU,UCC, UCA and UCG code for serine).
- Degeneration of genetic coding has several biological benefits. It enables, for instance, bacteria with vastly different DNA base compositions to specify virtually the same complement of enzymes and other proteins.
- Degeneration also provides a technique for decreasing the lethality of mutations.
7. Some codes act as start codons
- In the majority of organisms, the AUG codon is the start or initiation codon, meaning that the polypeptide chain begins with methionine (eukaryotes) or N-formylmethionine (prokaryotes) (prokaryotes).
- Methionyl or N-formylmethionyl-tRNA binds particularly to the start site of mRNA with an AUG initiation codon.
- In rare instances, GUG functions as an initiating codon, such as in bacterial protein production. GUG normally codes for valine; however, when the regular AUG codon is deleted, only GUG is used as an initiation codon.
8. Some codes act as stop codons
- Triple codons UAG, UAA, and UGA are the stop or termination codons for the chain. They do not code for any of the amino acids.
- These codons are not read by any tRNA molecules (via their anticodons), but are read by some specialised proteins, called release factors (e.g., RF-1, RF-2, RF-3 in prokaryotes and RF in eukaryotes) (e.g., RF-1, RF-2, RF-3 in prokaryotes and RF in eukaryotes).
- These codons are also called nonsense codons, since they do not designate any amino acid. The UAG was the first termination codon to be found by Sidney Brenner (1965). (1965).
- It was named amber in honour of a doctoral student named Bernstein (= the German term for ‘amber,’ and amber signifies brownish yellow) who helped identify a class of mutations.
- Apparently, the other two termination codons were also named after colours, such as ochre for UAA and opal or umber for UGA, in order to maintain consistency. (ochre indicates pale yellow or golden red, opal means milky white, and umber signifies brown)
- The presence of multiple stop codons may be a precautionary mechanism in case the first stop codon fails to work.
9. The code is universal
- The same genetic code is valid for all creatures, from bacteria to humans. Marshall, Caskey, and Nirenberg (1967) showed the universality of the code by showing that E.coli (bacterial), Xenopus laevis (amphibian), and guinea pig (mammal) amino acyl-tRNA utilise nearly the same code.
- Nirenberg has also suggested that the genetic code may have originated with the first bacteria three billion years ago, and that it has altered very little over the history of living species.
- Recently, inconsistencies between the universal genetic code and the mitochondrial genetic code have been revealed.

Codon and Anticodon
- The codon words of DNA are complementary to the mRNA code words (i.e., DNA codes run in the 3’→5′ direction whereas mRNA code words run in the 5’→3′ direction), as are the three bases composing the anticodon of tRNA (i.e., anticodon bases run in the 3’→5′ direction).
- Three bases of the anticodon pair with the mRNA on the ribosomes during the alignment of amino acids during protein synthesis (i.e., the translation of mRNA into proteins in the N2→COOH direction).
- For instance, one of the two mRNA and DNA code words for the amino acid phenylalanine is UUC, while the equivalent anticodon of tRNA is CAA.
- This suggests that the pairing of codons and anticodons is antiparallel. C pairs with G and U pairs with A in this instance.
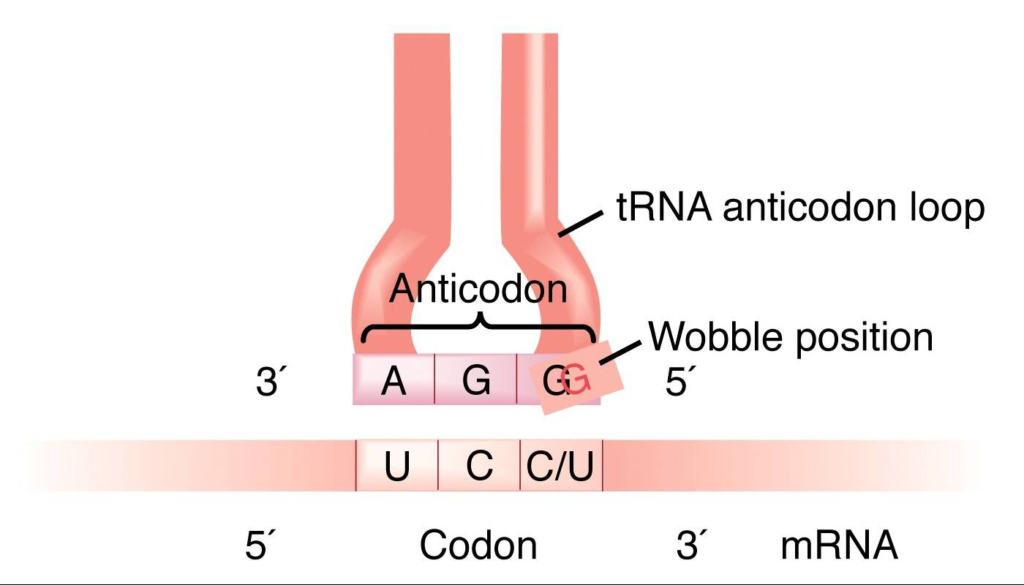
Wobble Hypothesis
- Crick (1966) presented the wobble hypothesis to explain the potential origin of codon degeneracy (wobble means to sway or move unsteadily).
- Given that there are 61 codons that specify amino acids, the cell must possess 61 tRNA molecules, each with a unique anticodon.
- The actual number of tRNA molecule types discovered is far fewer than 61. This suggests that tRNA anticodons read many codons on mRNA.
- For instance, yeast tRNAala with anticodon bases 5′ IGC 3′ (where I stands for inosine, a derivative of adenine or A) may bind to three codons in mRNA, including 5′ GCU 3′, 5’GCC3′, and 5′ GCA3′.
- Inosine is usually found as the 5′ base of the anticodon; when pairing with the base of the codons, it wobbles and can pair with U, C, or A of three different codons.
- Therefore, according to Crick’s wobble hypothesis, the base at the 5′ end of the anticodon is not as spatially constrained as the other two bases, allowing it to establish hydrogen bonds with any of the bases positioned at the 3′ end of a codon.

Leave a Comment Cancel reply
Save my name, email, and website in this browser for the next time I comment.

Adblocker detected! Please consider reading this notice.
We've detected that you are using AdBlock Plus or some other adblocking software which is preventing the page from fully loading.
We don't have any banner, Flash, animation, obnoxious sound, or popup ad. We do not implement these annoying types of ads!
We need money to operate the site, and almost all of it comes from our online advertising.
Please add Microbiologynote.com to your ad blocking whitelist or disable your adblocking software.
- Register / Log in
Wobble hypothesis tRNA wobble, Wobble position
A property of the genetic code in which codons that differ in the third position (wobble position) can specify the same tRNA/amino acid.
- Subscriber Services
- For Authors
- Publications
- Archaeology
- Art & Architecture
- Bilingual dictionaries
- Classical studies
- Encyclopedias
- English Dictionaries and Thesauri
- Language reference
- Linguistics
- Media studies
- Medicine and health
- Names studies
- Performing arts
- Science and technology
- Social sciences
- Society and culture
- Overview Pages
- Subject Reference
- English Dictionaries
- Bilingual Dictionaries
Recently viewed (0)
- Save Search

Michael Allaby
- Find at OUP.com
- Google Preview
- Share This Facebook LinkedIn Twitter
- wobble hypothesis in Oxford Dictionary of Biochemistry and Molecular Biology (2 ed.)
- wobble hypothesis in A Dictionary of Biomedicine
- wobble hypothesis in A Dictionary of Genetics (7 ed.)
- View overview page for this topic
Related Content
In this work, related overviews.
transfer RNA
genetic code
View all related overviews »
- Publishing Information
- General Links for this Work
- Preface to the Third Edition
- Plant Classification
- Kingdom Fungi
- The Universal Genetic Code
- The Geologic Time-Scale
- SI Units (Système International d'Unités)
- Previous Version
wobble hypothesis
A theory proposed to explain the partial degeneracy of the genetic code in that some t-RNA molecules can recognize more than one ... ...
Access to the complete content on Oxford Reference requires a subscription or purchase. Public users are able to search the site and view the abstracts and keywords for each book and chapter without a subscription.
Please subscribe or login to access full text content.
If you have purchased a print title that contains an access token, please see the token for information about how to register your code.
For questions on access or troubleshooting, please check our FAQs , and if you can''t find the answer there, please contact us .
- Oxford University Press
PRINTED FROM OXFORD REFERENCE (www.oxfordreference.com). (c) Copyright Oxford University Press, 2023. All Rights Reserved. Under the terms of the licence agreement, an individual user may print out a PDF of a single entry from a reference work in OR for personal use (for details see Privacy Policy and Legal Notice ).
date: 15 May 2024
- Cookie Policy
- Privacy Policy
- Legal Notice
- Accessibility
- [66.249.64.20|81.177.182.174]
- 81.177.182.174
Character limit 500 /500

An official website of the United States government
The .gov means it’s official. Federal government websites often end in .gov or .mil. Before sharing sensitive information, make sure you’re on a federal government site.
The site is secure. The https:// ensures that you are connecting to the official website and that any information you provide is encrypted and transmitted securely.
- Publications
- Account settings
Preview improvements coming to the PMC website in October 2024. Learn More or Try it out now .
- Advanced Search
- Journal List
- v.15(4-5); 2018
Celebrating wobble decoding: Half a century and still much is new
Paul f. agris.
a The RNA Institute, State University of New York, Albany, NY, USA
b Department of Biology, State University of New York, Albany, NY, USA
c Department of Chemistry, State University of New York, Albany, NY, USA
Emily R. Eruysal
Amithi narendran, ville y. p. väre, sweta vangaveti, srivathsan v. ranganathan.
A simple post-transcriptional modification of tRNA, deamination of adenosine to inosine at the first, or wobble, position of the anticodon, inspired Francis Crick's Wobble Hypothesis 50 years ago. Many more naturally-occurring modifications have been elucidated and continue to be discovered. The post-transcriptional modifications of tRNA's anticodon domain are the most diverse and chemically complex of any RNA modifications. Their contribution with regards to chemistry, structure and dynamics reveal individual and combined effects on tRNA function in recognition of cognate and wobble codons. As forecast by the Modified Wobble Hypothesis 25 years ago, some individual modifications at tRNA's wobble position have evolved to restrict codon recognition whereas others expand the tRNA's ability to read as many as four synonymous codons. Here, we review tRNA wobble codon recognition using specific examples of simple and complex modification chemistries that alter tRNA function. Understanding natural modifications has inspired evolutionary insights and possible innovation in protein synthesis.
Introduction
Translation of the Universal Genetic Code ( Fig. 1 ) into the amino acid sequence of proteins requires accurate and efficient decoding of mRNA (mRNA) on the ribosome by tRNA (tRNA). Sixty-one of 64 3-nucleoside codons of the mRNA, N1N2N3, are decoded in frame with a complementary sequence of the tRNA anticodon, N 34 N 35 N 36 . Three codons are recognized by protein factors and correspond to translation termination signals. Complementariness of base pairing between tRNA's anticodon and the mRNA codon, A•U, U•A, G•C and C•G where • denotes canonical Watson-Crick hydrogen bonding, does not explain how the 61 amino acid codons are decoded by far fewer tRNAs. Fifty years ago, Francis Crick published the Wobble Hypothesis. 1 At the time there was evidence to suggest that the first two positions, N1N2, of mRNA's 3-nucleoside codons were uniquely identified by the tRNA with some ambiguity in the third position. Crick offered the idea that uridine (U) at the first position of the anticodon, position 34 in tRNA ( Fig. 2 ), would base pair with guanosine (G) and that inosine (I), which at the time had only recently been found at position 34 in yeast tRNA Ala , 2 would base pair with uridine, cytidine (C) and adenosine (A). Thus, canonical, Watson-Crick base pairing with tRNA and mRNA on the ribosome was supplemented with non-canonical hydrogen bonding, ‘wobble’ base pairing, including that of a purine with another purine. Though conceding that the wobble position base pairing of two purines would widen the anticodon-codon double helix at the point of an I 34 ○A3 base pair where A3 is the third nucleoside of the codon and ○ denotes non-canonical hydrogen bonding, Crick excluded the possibility of a wobble position pyrimidine-pyrimidine base pairing. He argued that the narrowing of the helix would be too dramatic in comparison to the neighboring canonical purine-pyrimidine distances at the first and second positions.

The Universal Genetic Code. The Universal Genetic Code is shown in an atypical array to highlight those codons and their decoding by tRNAs discussed here. Fully degenerate codon boxes are shown in blue, split codon boxes in brown and stop codons in red.

The tRNA journey. The secondary structure of tRNA with its constituent domains marked in different colors (top left): Acceptor Stem (green); Dihydrouridine Stem and Loop, DSL (black); Anticodon Stem and Loop, ASL (red); Variable Stem and Loop, VSL (yellow); Thymidine Stem and Loop, TSL (blue). tRNA transcripts are processed by sizing and modification, some are spliced, before functioning in translation. Modification of tRNAs, particularly the anticodon stem and loop (ASL) domain at positions 32, 34, 37, 38 and 39, is an important step toward achieving functional chemistry and architecture. The wobble nucleoside, first of the anticodon, is position 34. Red and black highlights of mature tRNA after modification indicate the locations in the ASL where it is heavily modified.
The post-transcriptional modification of tRNA, the original soluble RNA, has been known for over 50 years pioneered by the extraction and characterization methods of Ross Hall in the 1960s. 3 At the time the Universal Genetic Code was unveiled, 4 there were several RNAs known to contain modified nucleosides, but their sequence locations and functions were a mystery before the evolution of RNA sequencing methods. By 1991, 25 years later, a sufficient number of modified nucleosides had been found to occupy the wobble position 34 of tRNA's anticodon that a Modified Wobble Hypothesis was advanced. 5 Biophysical and biochemical experiments had suggested and continue to support the principle that some position 34 modifications structure the architecture of the anticodon stem and loop (ASL) to counter wobble whereas other modifications shape the ASL to enable a tRNA to decode three or even four synonymous codons (codons differing only in the wobble position N3). 6 - 10
Of what importance are these ubiquitous and highly conserved anticodon modified nucleosides to the decoding of mRNA codons? In general, modified nucleosides of tRNA's anticodon stem and loop (ASL) domain are found within the stem at positions 27, 28, 31, 39, and 40 and at loop positions 32, 34, 35, 37, and 38 ( Fig. 2 ). 11 - 13 These ten nucleoside positions of the ASL are not simultaneously modified in any one tRNA, rather a set of 3–5 specific nucleosides are found to be modified in an individual tRNA. Many times in the sequencing of a tRNA species one finds that as many as three nucleoside positions of the seven residue loop are modified. Considering that the ASL of tRNA constitutes 17 of the molecule's ∼76 nucleosides and that the stem constitutes 5 base pairs, with 10 nucleosides, the modifications within the ASL loop represent a dense population of altered nucleoside chemistries. Often the modifications at wobble position 34 and 3′-adjacent to the anticodon at position 37 are the most chemically complex of all RNA modifications and are composed of hydrophobic aliphatic chains or aromatic substituents, or of highly hydrophilic or even charged functional groups participating in translational efficiency and fidelity. 11 Thus, posttranscriptional modifications of the ASL nucleosides emulate the chemistries of amino acid side chains. 14 Even what appears to be one of the simplest of modifications, such as the substitution of a sulfur atom for an oxygen, a thioketone for a carbonyl group, or the deamination of adenosine to inosine (I), takes on significance in the decoding of mRNA. The modification of wobble position U 34 to s 2 U 34 alters tRNA's ability to wobble to G3; 5 the modification of C 32 to s 2 C 32 negates the ability of tRNAs with I 34 to decode A3 of the codon. 6
The importance of being modified
The high percentage of anticodon modification and their composite chemistries provides different functionalities to tRNA in its role in decoding mRNA ( Fig. 3 ). Individual anticodon domain modified nucleosides are identity determinants for protein recognition, particularly aminoacylation, 15 - 17 increase accuracy and efficiency in codon recognition, 18 - 24 and pre-structure the ASL for translation. 9 , 25 - 30 Each of the modified nucleosides contribute distinct chemistries, nucleoside conformations and dynamics, and their contributions to decoding have been studied extensively over decades and most recently reviewed. 20 , 22 , 24 , 28 , 31 - 38 However, there is significant evidence that a combination of two or three anticodon domain modifications play a synergistic role in tRNA function where modification of a wobble position U is crucial. 9 , 20 , 24 , 39 - 46 Today, we know that certain modifications of U 34 enable expansion of codon from NNA/G recognition to synonymous codons ending in pyrimidines, N1-N2-Pyr, where N is any of the 4 nucleosides and Pyr is either U or C. 29 Yet, the anticodon domain of some tRNA species lack modification and can be totally devoid of modified nucleosides. Bacteriophage T4 tRNA Gly is an early example of a tRNA lacking modification. 47 The unmodified U 34 nucleosides of native mitochondrial alanine, leucine, threonine and valine tRNA species in vivo , and that of a totally unmodified tRNA transcript in vitro were shown to read codons ending in U3 and C3, as well as A3 and G3. 48 , 49 To understand the forces that maintained and propagated tRNAs' anticodon domain modifications throughout all life, first we will discuss the limited number of examples revealed over many years to have unmodified Us and As at wobble position 34, and the influence of position 32 nucleosides on decoding.

Modifications present in tRNA's anticodon stem and loop domain (ASL) featured in the text and displayed in their neutral state. A. Modified purines, adenosine (A) and guanosine (G). B. Modified pyrimidines, uridine (U) and cytidine (C). R = ribose.
Unmodified wobble position 34 and the importance of position 32
Bacteria, archaea and eukaryotes have some 40 tRNA species decoding the Universal Genetic Code. However, many times we find that in mammalian and yeast mitochondria, in chloroplasts, and in Mycoplasma ssp only a single tRNA species decodes all 4 codons of a fully degenerate codon box of the Universal codes ( Fig. 1 ). 49 - 57 Wobble codon recognition is at its extreme in these circumstances and has been referred to as ‘superwobble’. 58 An unmodified U in tRNA's wobble position 34 facilitates the use of far fewer tRNAs in organelles than the over 40 cytoplasmic species typically reading the 61 amino acid codes. Fewer tRNAs is certainly an advantage for the small genomes of the organelles and the minimal single cell organism, mycoplasma. An unmodified U 34 also abrogates the need for the extensive array of genes encoding the modified nucleoside pathways of enzymes and substrates required of the simplest to the most complex modifications of U 34 . 11 However, the superwobble reading of codons compromises translational efficiency. 58
An unmodified U 34 seems to be particularly efficient for the organellar tRNA Gly species. The codons for the amino acid glycine, GGA, GGG, GGU or GGC, are found in a 4-fold degenerate codon box, whose first two nucleosides are the same ( Fig. 1 ). They are read by as many as three different tRNA isoacceptors in bacteria with cognate and wobble anticodons. Early in the study of glycine tRNAs, the Escherichia coli tRNA Gly isoacceptors were grouped into 3 subspecies based on a chromatographic separation: tRNA Gly1 CCC , tRNA Gly2 UCC , and tRNA Gly3 GCC . 59 In an E. coli in which there were multiple copies of suppressors increasing the levels of wild-type tRNA Gly1 CCC , −1 translational frameshifting occurred at the 5′-GGG-3′ codon allowing the near-cognate tRNA to read GGA codons. 60 Surprisingly, experiments with E. coli tRNA Gly2 UCC demonstrated that the unmodified UCC anticodon discriminates among the four glycine codons depending on the nucleoside in position 32, an unmodified U 32 or C 32 ( Fig. 4A ). Thus, the unmodified UCC anticodon reads GGA and wobbles to GGG, but does not recognize GGU and GGC. 61

Sequences of unmodified ASL of tRNA Gly . A. ASLs of E. coli tRNA Gly1 CCC , tRNA Gly2 UCC , and tRNA Gly3 GCC indicating the changes in nucleoside position 32 and 34 (marked in red) from the tRNA Gly1 sequence. All 3 E. coli tRNA Gly have the same nucleoside at position 32 (U 32 ). The Mycoplasma mycoides tRNA Gly UCC differs from the E. coli tRNA Gly as a result of a cytidine at position 32 as well as the presence of inverted base pairs in the anticodon stem at positions 27•43 and 28•42. B. Three Dimensional Structure of the tRNA Gly GCC and tRNA Gly UCC ASLs of B. subtilis . The interactions between the U 32 •A 38 and C 32 ○A 38 nucleosides (PDB ID: 2LBJ and 2LBL) are shown. Both ASLs lack the sharp U-turn characteristic of ASLs of other tRNAs. Nucleosides of the anticodon are labeled 34 to 36.
Although the anticodon UCC could discriminate efficiently with a U in position 32, it loses its ability to differentiate when substituted with a C 32 as was also true for M. mycoides glycine tRNA. In wild type M. mycoides tRNA Gly , the anticodon UCC failed to discriminate between the glycine codons with a position 32 cytidine, but when changed to a U it acted like E. coli tRNAs and discriminated between the four glycine codons. 62 In M. mycoides , the single tRNA Gly UCC that decodes all four glycine codons is devoid of anticodon modifications and has a C 32 ○A 38 mismatch which leads to decreased fidelity in vivo ( Fig. 4 ). 63 The tRNA Gly UCC transcript without any modifications reads all 4 codons in vitro . 48 Ribosome binding experiments with U 32 /C 32 mutants of tRNA Gly showed an increased affinity of the C 32 mutant to the cognate codon and to codons with third position mismatches in the ribosome's A-site. 64 The rate of dissociation of the U 32 -containing tRNA Gly from the near-cognate GGC, GGA and GGU codons was much more rapid - 12-fold faster than from the GGG cognate codon - and stabilized the binding of tRNA to codons with third position mismatches. 64 In contrast, the mutated tRNA Gly1 with C 32 dissociated much more slowly from near-cognate codons. Analysis of the UV-thermal denaturation (melting) curves of the anticodon stem and loop domains demonstrated that tRNA Gly UCC with a protonated C 32 ○A + 38 non-canonical base pair melted at a temperature 10 °C lower than tRNA Gly GCC or np-tRNA Gly UCC , used exclusively for non-protein cell wall synthesis in Staphylococcus species, which exhibited a T m of 70 °C. 65 Although the sequences of these tRNA differed from one another, none of their solution structures formed the classical U-turn motif seen in other tRNA anticodon loops ( Fig. 4B ). tRNA molecules fulfill different functional roles by interacting with other cellular molecules, and the structural variations of the glycine ASL of S. aureus could contribute to its functional diversity. tRNA Gly GCC , without any base modification participates in transcriptional regulation and transcription. The tRNA Gly UCC which more often contains a U 34 modification except in some organisms including M. mycoides , participates in translation. The np-tRNA Gly UCC contains no base modification and participates in cell wall synthesis. 65 The presence of pyrimidine at position 37, along with a reduced affinity for EF-Tu, could limit their involvement in transcription and translation. Modifications of U 34 could increase or decrease the ability to wobble, thus enhancing discrimination. A more dynamic ASL tRNA Gly GCC was observed in the presence of multivalent cations, whereas tRNA Gly UCC and np-tRNA Gly UCC were more structurally ordered in their presence. A more dynamic loop structure would therefore, better accommodate the different functional roles of an unmodified tRNA Gly in protein translation, in tRNA-dependent gene regulation, and cell wall biosynthesis. 65
The nucleoside at position 32 of the tRNA's anticodon loop is recognized as important for translation, though the position 32 nucleoside is remote from the anticodon. The intraloop hydrogen bonding between the nucleosides at 32 and 38 strongly influences the affinity of tRNA to the A-site. 64 tRNA species other than those for glycine also have an unmodified wobble position U 34 , and the nature of the N 32 ○N 38 intraloop hydrogen bonding in these tRNAs does appear to regulate expansion/discrimination of codon reading. The A 32 •U 38 interaction, highly conserved in tRNA Ala and also seen in tRNA Pro , decreases the tRNA affinity to the ribosomal A-site compared with other N 32 ○N 38 intraloop hydrogen bonding nucleosides. 66 Five Mycoplasma capricolum tRNA species with the unmodified anticodon UNN and decoding for five different amino acids have anticodon loop sequences with C 32 ○A 38 . In addition, tRNA Ala UGC has an intraloop interaction of C 32 ○C 38 . Because these 6 tRNA species have a C 32 and an unmodified U 34 , in theory they could efficiently read all four synonymous codons for leucine, valine, proline, alanine, glycine and serine. 67 Although tRNA Thr UGU was shown to translate the codons ACU, ACA and ACG, it was inefficient in reading the codon ACC.
Interestingly, the sufD42 mutant of E. coli tRNA Gly1 , encoded by glyU, is a derivative of tRNA Gly CCC with an extra C in the anticodon loop and contains no modification in the anticodon loop. 68 This mutant is considered dominant and contains four bases, 5′-CCCC-3′ that make up the anticodon and suppress +1 frameshift mutants with an extra G inserted into a GGN codon (GGGN). The quadruplet translocation theory is used to deduce the pairing of these four cytidines with four bases of the codon in the A site. The tRNA anticodon sequence has been suggested to act as a molecular ruler which determines the codon size during translation, within certain limits, 69 thereby restoring some ribosomes to the wild type frame. Thus, the nature of the N 32 ○N 38 base interaction affects the binding of the anticodon to the codon suggesting that the intraloop hydrogen bonding alters the conformation and dynamics of the anticodon stem and loop domain within the ribosomal complex. 62 , 70
Very few unmodified, wobble position A 34 have been found in tRNA sequences: a yeast mitochondrial tRNA Arg ACG 71 and Mycoplasma tRNA Thr AGU . 50 , 56 , 72 In all domains of life, A 34 of the tRNA transcript is almost always deaminated to form inosine at the wobble position (I 34 ). The modification to I 34 expands the coding capacity to read the bases U, C and A. In Mycoplasma the unmodified A 34 of tRNA Thr AGU efficiently translates the ACC codon, 67 but mutants of E. coli tRNA Ser and tRNA Gly with A 34 could only weakly read UCC and GCC in vitro , respectively. 73 , 74 In Salmonella typhimurium , tRNA Pro GGG is the only tRNA that reads the CCC codon. With G 34 replaced by an unmodified A, a mutant with no cognate codon for CCC, grew normally. 75 The mutant tRNA Pro AGG efficiently read the CCC codon similarly to its wild-type counterpart with a GGG. It formed a wobble base pair using a protonated A with the third position C in mRNA. Similarly, a mutant tRNA Gly1 with a UCC to ACC mutation containing an unmodified A 34 , lost its ability to discriminate between the third position nucleosides of the glycine codons. 73 The possibility of a purine○purine base pair being more stable than a pyrimidine○pyrimidine pair, 76 as well as the two out of three model 77 was suggested to explain the non-discrimination by A in the wobble position. 73 The presence of A in the wobble position could change the conformation of the anticodon by preventing hydrogen bonding between U 33 and the phosphate of nucleoside 36, 78 which stabilizes the U-turn conformation of the ASL. The rare occurrence of an unmodified A 34 is supported by the hypothesis that a wobble position A cannot discriminate and ensure translational fidelity of tRNAs reading split box codons. In contrast, the presence of A 34 would be advantageous in the reading of fully degenerate synonymous codons where there is a lack of discrimination and in cases where only a single tRNA exists for the reading all four codons. 73
In addition to their primary role of translation, some tRNAs have other functions including regulation of gene expression, bacterial cell wall synthesis, viral replication, antibiotic biosynthesis and suppression of alternative splicing. 64 , 65 In Bacillus subtilis and many other Gram-positive bacteria, tRNA molecules regulate gene expression by the tRNA dependent, ‘T-box’, mechanism of transcription attenuation to maintain a balanced pool of aminoacyl-tRNAs that is essential for cell viability. 79 , 80 The tRNA ligand for the T-box mechanism regulating the expression of glycyl-tRNA synthetase is tRNA Gly UCC with an unmodified U 34 . A Rho-independent, terminator helix in the 5′UTR of the leader mRNA of the glyQS operon for glycyl-tRNA synthetase prevents the operational binding of an aminoacylated glycyl-tRNA Gly UCC . 81 Conversely, the anticodon of an uncharged tRNA interacts with a loop, the Specifier Loop, containing the complementary codon and the tRNA's 3′-terminal CCA hydrogen bonds to an anti-terminator helix, re-conformed from the terminator helix. These interactions as well as others between the tRNA and the mRNA stabilize the anti-terminator conformation of the 5′UTR and allow transcription to proceed downstream through the coding sequence. Thus, the unacylated tRNA Gly UCC is similar to the much smaller metabolic products that affect the riboswitch mechanisms controlling gene expression; the 5′UTR undergoes a conformational change with the binding of the ligand. 82 The tRNA Gly UCC is also predicted to bind to the 5′-GGA-3′ Specifier codon in Bacillus and Staphylococcus species glycyl T-box riboswitches. 83
A third glycine tRNA (UCC) without a modified U 34 has been identified in Staphylococcus species as participating in cell wall biosynthesis, but not in protein translation, and was termed non-proteinogenic (np-tRNA Gly ). 84 - 87 These np-tRNA have an unmodified U 34 nucleoside and a cytidine rather than a purine at position 37. 88 In Thermus thermophiles , the np-tRNA Gly species are found to have reduced affinity for the elongation factor Tu (EF-Tu) due to base substitutions of A 51 -U 63 for G 51 -C 63 in the base of the T-stem, thus decreasing their involvement in ribosomal protein synthesis. 65 These weak EF-Tu binders could act as glycine donors in forming essential pentaglycine bridges which stabilize the staphylococcal cell wall. 85 , 89 - 91
The biosynthesis of peptidoglycan in S. aureus involves two uridine nucleotide substrates, UDP-MurNAc-pentapeptide and UDP-GlcNAc (N-acetyl glucosamine), which combine to form a lipid intermediate GlcNAc-MurNAC(pentapeptide)-P-P-lipid. The lipid intermediate gets further modified by amidation of the α-carboxylic group of glutamic acid and the addition of a pentaglycine chain to the ε-amino group of lysine. The weak EF-Tu binding glycyl tRNAs serve as intermediates in these reactions to form the pentaglycine bridges, that stabilize the peptidoglycan chains and are essential for cell viability. These short peptide bridges are synthesized in a non-ribosome catalyzed peptidyl transferase reaction, which uses the charged glycyl-tRNA, 1 np-tRNA Gly and a ‘pseudo’-tRNA Gly UCC as substrates. 65 , 85 , 92 Glycine and serine act as substrates that are successively added to form small peptide bridges which are catalyzed by a family of non-ribosomal peptidyl transferases known as FEM-XAB (Factors Essential for Methicillin Resistance) - mediated cell wall synthesis. 93 The incomplete formation of these interpeptide bridges can lead to increased antibiotic susceptibility or lethality. 85
The wobble hypothesis and the modulation of inosine wobbling
Though there are instances of unmodified nucleosides at tRNA's wobble position 34 as described, more often than not U 34 is post-transcriptionally modified and A 34 is deaminated to inosine. Using specific modifications of U and the modulation of I reading A, U and C, we illustrate here the importance of wobble position 34 modifications to tRNAs' accuracy and efficiency of translation. The modification of adenosine to inosine was first recognized by Francis Crick for enabling tRNA recognition of synonymous codons. 1 Inosine ( Fig. 3 ) results from the deamination of adenosine, a transformation that is facilitated by the adenosine deaminase (ADAR) family of enzymes that act on RNA. 94 Although inosine is a marker of damage or mutation in DNA, the presence of this very same modified nucleoside is considered to be essential in various RNAs. 95 Inosine plays a vital role in the function of tRNA, in particular. As the first recorded nucleoside modification within the sequence of an anticodon, 2 Crick introduced inosine in his 1966 Wobble Hypothesis. 1 While the first two bases of the codon undergo traditional base-pairing without exception, 96 Crick proposed the potential for non-canonical base pairs between the first base of the anticodon (“wobble” position 34) and the third base of the codon, U 34 ○G3, or I 34 ○A3/U3/C3 ( Fig. 5 ). 1 This flexibility of the genetic code is not without limitations; however, in accordance with this hypothesis, a given tRNA isoacceptor may recognize multiple codons, thus explaining the degeneracy of the genetic code.

Canonical and wobble base pairing of tRNA to mRNA. A. Canonical A•U and G•C base pairs. B. Wobble U 34 ○G3, I 34 ○A3, I 34 ○C3, and I 34 ○U3 base pairs. G 34 ○U3 pairings are virtually nonexistent; therefore, the pairing is not shown. The arrows point away from the hydrogen bond donor and toward the hydrogen bond acceptor.
The Wobble Hypothesis states that position 34 inosine may base pair with uridine, cytidine, and adenosine. The ability of inosine at the wobble position to promote the reading of multiple codons, in some cases, proves essential to survival. The heterodimeric enzyme consisting of the Tad2p and Tad3p subunits of Saccharomyces cerevisiae catalyzes the deamination of adenosine to inosine on tRNA. 97 A strain of Schizosaccharomyces pombe containing a mutant tad3–1 , the homolog of TAD3 , experienced temperature-dependent arrested growth at Gap 1 and Gap 2 of the cell cycle. 98 The S. pombe genome utilizes only 3 tRNA Ala isoacceptors for the 4 alanine codons GCU, GCC, GCG, and GCA: tRNA Ala IGC , tRNA Ala CGC , and tRNA Ala UGC . According to the wobble rules, tRNA Ala IGC must be responsible for decoding of the GCC codon. The otherwise unmodified tRNA Ala AGC would be able to decode its complementary GCU codon but could not wobble to GCC, inhibiting the translation of gene products vital to the G 1 /S and G 2 /M transitions of the cell cycle. 98
A tRNA Arg IGC may be modified from A 34 to I 34 yet still be unable to decode A3. The E. coli tRNA Arg1 ICG and tRNA Arg2 ICG isoacceptors should bind and effectively decode the CGU and CGC codons, and even the CGA codon. These wobble pairings were confirmed experimentally, as the singly modified anticodon stem and loop of tRNA Arg ICG , ASL Arg ICG , is able to bind the CGU, CGC, and CGA codons within the ribosomal A-site. 39 In fact, tRNA Arg1 ICG and tRNA Arg2 ICG are the only isoacceptors available to recognize the 3 aforementioned codons. Furthermore, adenosine must be modified to inosine at position 34 of the tRNA Arg1,2 ICG isoacceptors in order for wobble pairing to occur. Unmodified ASL Arg1,2 ACG is able to bind its cognate codon, CGU, but unable to bind CGC or CGA as expected, as Crick did not explicitly delineate any wobble capabilities of position-34 adenosine. 1 , 39
The tRNA Arg1,2 ICG decoding of the CGA codon is relatively inefficient compared with translation of the CGU. 99 , 100 Although the energy barrier for I○A base-pair formation is greater than those of I○C and I○U, the increased distance between the N-glycosyl bonds of I 34 and A3 required to accommodate the purine○purine wobble pair can be achieved when the nucleosides adopt an I anti ○A anti conformation. 1 , 44 , 101 Additional modifications at position 32 and 37 of the ASL Arg ICG may further contribute to the difficulties of I○A wobble pairing. The E. coli tRNA Arg1 ICG species contains the naturally-occurring 2-thiocytidine at position 32 (s 2 C 32 ) and 2-methyladenosine at position 37 (m 2 A 37 ). The thrice modified ASL construct could not be synthesized, but the doubly modified ASL Arg ICG -s 2 C 32 was unable to bind the CGA codon within the ribosomal A-site. 39 Here, the complete wobble capabilities of inosine do not apply due to the restrictive effects of the s 2 C 32 modification with respect to the otherwise feasible I○A wobble pair.
The tRNA Arg2 ICG species lacks the s 2 C 32 modification but does contain the m 2 A 37 modification. As evidenced by the aforementioned ribosomal binding study, m 2 A 37 prohibits I○A pairing. 39 However, reading of the CGA codon defaults to tRNA Arg2 ICG , as the inclusion of the s 2 C 32 modification disqualifies the tRNA Arg1 ICG from decoding of the CGA codon in vivo . Yet the E. coli genome still must compensate for the overall poor capacity of the tRNA Arg ICG isoacceptors to wobble to the CGA codon by biasing codon usage. The inclusion of CGA codons in mRNA transcripts increases energetic costs and decreases the efficiency of translation. The prevalence of the CGU, CGC, and CGA codons is heavily biased against the CGA codon and in favor of the CGU and CGC codons. 102 , 103
The modified wobble hypothesis and decoding at position 34
For many years, Crick's Wobble Hypothesis appeared to sufficiently explain the function of modified nucleosides at tRNA's wobble position without the need for alteration. However, the discovery of numerous new modifications, a large number of which are found exclusively at the wobble position, led to the development of a modified wobble hypothesis. 5 The first base of the anticodon is so often modified to either expand or restrict the binding abilities of the wobble nucleoside, therefore enabling the specific recognition of cognate and synonymous codons. As such, near-cognate codons can be selected against, or the recognition of multiple codons can be made feasible with various chemical moieties introduced onto the wobble base. 5 There are several examples of both expansion as well as restriction of codon recognition which are presented with a focus on the mechanistic details of both expansion and restriction of recognition, and the many factors that must come to play to make either possible.
Expansion of codon recognition through modified nucleosides pre-structuring of the ASL
The high freedom of rotation coupled with the limited chemical variation of the four major nucleosides enables RNAs to adopt multiple conformations with comparable stability yet limited chemistry. Modifications add to the chemistry and can either limit or expand the number of available conformations, thereby influencing the structure toward a more specific architecture or provide dynamics important to translational effectiveness. There are six amino acids represented by 4-fold degenerate codon boxes which include alanine, glycine, proline, serine, threonine, and valine, and are of particular interest in U 34 modification. As with S. pombe and the GCN alanine codon box, Salmonella enterica utilizes 3 distinct species of tRNA Pro for recognition of the entire CCN codon box: tRNA Pro CGG , tRNA Pro GGG , and tRNA Pro UGG . Deletion of the genes for tRNA Pro CGG and tRNA Pro GGG simultaneously is not lethal to S. enterica . 104 Only tRNA Pro UGG proves necessary for viability of the organism by surpassing its expected decoding capabilities due to the presence of the 5-oxyacetic acid modification (cmo 5 ) on position-34 uridine in the ASL. The cmo 5 U 34 modification is present in one species of each of the tRNA isoacceptors for alanine, proline, serine, threonine, and valine. Similarly, tRNA Val UAC -cmo 5 and tRNA Ala UGC -cmo 5 partially rescue the E. coli growth phenotype caused by knockout of the 2 tRNA Val GAC isoacceptors and the tRNA Ala GGC isoacceptor, respectively, indicating the ability of tRNA containing the cmo 5 U 34 modification to decode an entire codon box, albeit somewhat inefficiently. 105 The presence of the cmo 5 U 34 modification facilitates pre-structuring of the ASL Val UAC that enhances its ability to bind near cognate codons. When binding each of the GUN valine codons, the ribose of cmo 5 U 34 of the modified ASL Val UAC assumes the C3′- endo conformation. 29 Prevalent within A-type helical regions of tRNA, the C3′- endo sugar pucker is synonymous with stability and rigidity. 106 - 108 The existence of a hydrogen bond between the 2′-OH of the almost invariant U 33 of the ASL and O5 of the cmo 5 modification further constrains cmo 5 U 34 . 29 This intramolecular hydrogen bond in particular expands the ability of ASL Val UAC -cmo 5 U 34 to decode codons ending in U and C, pre-structuring the ASL such that the entropic cost of a pyrimidine○pyrimidine base pair, initially believed to be unfavorably short, is surpassed. 1 , 29
The cmo 5 modification has surprising implications for the U 34 •G3 wobble pair. Although this pairing was originally predicted without regard for the potential of modified uridines, the uridine at position 34 of the ASL must be modified in order for the U 34 •G3 pairing to occur. 18 , 105 Rather than enhancing the ability of U 34 to wobble to G3, cmo 5 merely enables the interaction. In a second divergence from the original Wobble Hypothesis, the modified cmo 5 U 34 •G3 does not adopt the predicted wobble geometry with two hydrogen bonds between the two bases. The cmo 5 U 34 instead forms three hydrogen bonds with G3 as in the traditional Watson-Crick geometry ( Fig. 6 ). 29 The steric and electronic properties of the cmo 5 modification are thought to promote the enol form of cmo 5 U 34 , thus allowing for the Watson-Crick geometry of the cmo 5 U 34 •G3 base pair reminiscent of a C•G base pair. 29 , 109

Watson-Crick geometry of the cmo 5 U 34 ○G3 base pair. A. The predicted geometry of the unmodified U 34 ○G3 base pair containing two hydrogen bonds. B. The observed geometry of cmo 5 U 34 ○G3 base pair containing three hydrogen bonds. For the cmo 5 U 34 ○G3 to resemble a C•G base pair, the cmo 5 modification is proposed to facilitate formation of the enol tautomer. The arrows point away from the hydrogen bond donor and toward the hydrogen bond acceptor.
Additionally, E. coli tRNA Val UAC containing the cmo 5 U 34 modification is also methylated at the position-37 adenosine to yield N 6 -methyladenosine (m 6 A). As seen with tRNA Arg1 ICG -s 2 C 32 ;m 2 A 37 and tRNA Arg2 ICG -m 2 A 37 , it is not uncommon for the ASL of a given tRNA to contain multiple modified nucleosides. Modifications written onto N6 of the universal purine located at position 37 of the ASL are proposed to interfere with intraloop hydrogen bonding and to enhance base-stacking, thereby promoting an open loop structure within the ASL. 18 The open loop structure of the ASL is a prerequisite for anticodon-codon binding within the ribosomal complex. 110 ASL Val UAC lacking both the cmo 5 U 34 and m 6 A modifications does not appreciably bind the GUU, GUC, or GUG codons within the 30S ribosomal subunit. 111
While G is able to base pair with both C as well as U, the interaction with the latter is thermodynamically less stable. As such, tRNAs coding for aspartate, asparagine, histidine, and tyrosine commonly contain 7-(((4,5-cis-dihydroxy-2-cyclopenten-1-yl)amino)methyl)-7-deazaguanine, queuosine or Q ( Fig. 3A ), at the wobble position, which enables recognition of codons ending in either C or U without favoring one or the other. 112 This is true even for the more simplistic and somewhat less commonly modified mitochondrial tRNAs, implying an evolutionary necessity for enhanced codon recognition that Q provides. 113 Q is synthesized by bacteria, and eukaryotes acquire Q either as a nutrient or from the intestinal flora as quenine, the free base form of Q, which is then directly inserted into the pertinent tRNAs through substitution of the base quenine. 114 Interestingly, the E. coli tRNA Tyr contains pseudouridine at position 35, the second position of the anticodon and adjacent to Q, ms 2 i 6 A at position 37, and Ψ at position 39. Loss of Ψ at the rarely modified position 35 compromises translational fidelity in vitro , implying that some ASLs need even more than three modified nucleosides for accurate function. 115 Like many other large modifications, Q causes a steric effect in the ASL, influencing the structure of the loop. The effect is largely mediated by hydrogen bonding within the ASL, which has been shown to maintain the structure of the ASL in silico , thus pre-structuring the ASL for codon recognition in the ribosome. The electrostatic potential of Q compared with G is important. Due to the intramolecular hydrogen bond between O6 and the quaternary amine of the aminomethyl side chain, the electronegativity of O6 in Q is reduced compared with O6 in G. In G, O6 acts as a hydrogen bond acceptor, however, this is not the case for Q. The two amines act as hydrogen bond donors in both species. The side-chain of Q can also engage in hydrogen bonding with the backbone of U 33 , which in turn enhances the U 33 ○C 36 interaction, stabilizing the ASL conformation. This same effect is absent or reduced if Q is replaced with G. 112 The pre-structuring is further enhanced by the large modification wyosine, (3,4-dihydro-4,6-dimethyl-3-β-D-ribofuranosyl-9 H -imidazo[1,2-α]purin-9-one), imG, also derived from G and found at position 37 along with other derivatives often paramount to tRNA structure and function. 112
The high degree of hydrophobicity of wyosine ( Fig. 3A ) and its derivatives enhances the nucleoside's ability to form hydrophobic interactions such as base stacking, which enhances codon binding stability. The bulky residue prevents the intraloop hydrogen bond between residues 32 and 37, thus promoting proper pre-structuring of the ASL. 116 , 117 In mitochondrial tRNAs, imG 37 or its derivatives are commonly replaced by i 6 A 37 or a derivative thereof, or by a methylated purine, therefore suggesting the evolution of an alternative strategy for establishing the same functional properties by using different chemical moieties in RNA modifications. 113 , 116
The nearly universal modification at position 37 that is required for decoding codons beginning with A, N 6-threonylcarbamoyladenosine, t 6 A 37 , may be further modified to 2-methylthio- N 6-threonylcarbamoyladenosine, ms 2 t 6 A 37 . 118 Interestingly, a cyclic derivative of t 6 A, ct 6 A, has also been shown to exist, functioning in a manner analogous to the more studied t 6 A. 118 , 119 Hydrogen bonding between N1 and N11 of t 6 A lead to a pseudocyclic conformation that enhances the rigidity of the conformation. 120 The same hydrogen bonding exists in ct 6 A, but a further isomerization occurs as well, where C10 and C13 are linked via an ether, thus forming a oxazolidine. 118 - 120 The C14 alcohol of ct 6 A can hydrogen bond with the N7 of the first codon, an A, increasing the stabilization that enhanced stacking alone provides. 118 , 120 , 121
Restriction of codon recognition and pre-structuring of the ASL
RNAs are highly flexible molecules with each base containing several bonds about which free rotation can take place. The flexibility and the possibility of not only Watson-Crick base pairing but also wobble or Hoogsteen interactions provide suitable conditions for multiple structures to form. Certain modifications restrict intraloop base pairing interactions, thus conforming the architecture to one suitable for ribosomal decoding. Restriction of codon recognition may become necessary in cases where an unmodified nucleoside could base pair with a near-cognate codon, therefore causing misreading of the codon. U, for instance, can base pair with all of the 4 major nucleosides with a strong preference to purines. 18 As such, modification can provide an enhanced specificity. As an example, isoleucine shares a codon box with methionine where AUA, AUC, and AUU code for isoleucine, and AUG codes for methionine, with the exception of mitochondria where AUA also codes for methionine. 6 , 113 , 122 Two codons can be read by tRNA Ile GAU , but the AUA codon requires a different isoacceptor that would also not read AUG as isoleucine. Eukaryotes commonly have a tRNA Ile IAU which can read all three codons, but not AUG. 122 The genomes of bacteria and some eukaryotic organelles encode a tRNA Ile GAU and tRNA Ile LAU where L is lysidine (k 2 C). 123 Modification of C to k 2 C ( Fig. 3B ) effectively changes the base pairing capabilities of the nucleoside, causing a switch in preference from G to A for tRNA Ile . The modification is required to prevent the recognition of the near-cognate AUG codon ( Fig. 7 ). 7 Lysidine is a modified cytidine containing a lysine residue in lieu of O2 at the wobble position. The modification lysidine is the recognition determinant for the amino acid specificity in isoleucyl-tRNA synthetase (IleRS) aminoacylation of tRNA Ile LAU and not tRNA Met CAU . An intriguing minor tRNA Ile ΨAΨ in yeast contains pseudouridine at position 34 instead of lysidine, 121 and many archeal species utilize agmatidine [N-(4-carbamimidamidobutyl)-4-imino-1-(b-D-ribofuranosyl)-1,4-dihydro-2-pyrimidinamine] ( Fig. 3B ) for decoding of the AUA isoleucine codon. Thus, there have been multiple and convergent evolutionary paths to accomplish specificity in decoding through restriction of codon recognition with the help of modified nucleosides. 122

Modifications influence protein and codon recognition of tRNA through altered hydrogen bonding. R represents the ribose sugar. Hydrogen bonding is indicated by arrows. A. Nucleosides commonly found at position 34 of tRNA Ile . All 4 bases are capable of recognition by isoleucyl-tRNA synthetase, IleRS. B. Two possible tautomers of lysidine. Structure on the right can be recognized by IleRS, and could putatively recognize A as shown. C. Possible tautomers of mnm 5 s 2 U 34 of E. coli tRNA Lys UUU . Likely interactions with A and G are shown.
Tautomerism: A chemical mechanism by which modified nucleosides restrict codon recognition
Tautomers and rare ionic forms of nucleosides have been shown to exist at low populations in vitro ; 124 it is commonly postulated that the existence of tautomers of modified nucleosides at the ribosome's decoding site has a strong effect on codon recognition, either preventing or enabling it. 28 The nature of the C5 modification affects the tautomerism of the uridine and 2-thio modified uridine 125 producing either an expanded codon recognition, or one which is restricted. Restriction of codon recognition is a common mechanism for fidelity when a tRNA anticodon can mistakably base pair with a near-cognate codon in a shared codon box. In contrast, expansion of codon recognition by tRNA is often required when an interaction of an unmodified nucleoside would either lack the base pairing ability, thermodynamic stability, or the required structural orientation to achieve accuracy and efficiency of translation. 18
Both lysidine and agmatidine can exist in several tautomeric forms. The exact tautomeric form through which cognate codon recognition can occur still needs to be elucidated. 122 However, it is a tautomer of lysidine that disallows recognition of the near-cognate AUG codon. The modification contains a secondary amine in place of oxygen at the two position of C, thus switching a hydrogen bond acceptor into a donor. The two tautomers ( Fig. 7B ) have different base pairing capabilities due to distinct local electron densities. Interestingly, neither tautomer is ideal for base pairing with G, but the non-aromatic tautomer is capable of recognizing A; the primary amine at position 4 on the ring is switched into an imine, thereby enabling it to function as a hydrogen bond acceptor. Similarly, N3 of the base becomes a hydrogen bond donor, thus enabling base pairing with A ( Fig. 7B ). 126 Agmatidine forms a similar tautomer as lysidine. 122 , 126 The hydrogen bonding capability of modified nucleosides commonly found at the wobble position of tRNA Ile illustrates how small changes in the chemistry of a single nucleoside can have broad and highly specific effects, in this case altering codon recognition and specificity while simultaneously preserving aaRS recognition. 121
Tautomers of modified nucleosides expand reading of synonymous codons
U•A base pairs are thermodynamically much weaker than C•G base pairs. In tRNAs such as tRNA Lys UUU this problem is exacerbated, as all anticodon to codon base pairs are U•A pairs. Furthermore, in the absence of modifications, the U-rich ASL cannot effectively form stacking interactions to stabilize its structure nor negate intraloop hydrogen bonding that effectively condenses the size of the ASL. 117 The nearly ubiquitously modified, invariable purine at position 37 can alleviate the thermodynamic penalty of a pyrimidine rich anticodon. In tRNA Lys UUU the purine at position 37 is modified to N 6 -threonylcarbamoyladenosine, t 6 A or a derivative thereof such as 2-methylthio- N 6 -threonylcarbamoyladenosine, ms 2 t 6 A 37 found in mammalian tRNA Lys3 UUU . Additionally, Ψ at position 39 is important for function. 127 The wobble position is extensively modified with an xm 5 s 2 U type of modification, illustrating the importance of having three modified nucleosides in the anticodon loop for at least this tRNA. 28 , 128
Lysine is coded by AAA and AAG codons. In many organisms, as well as in organelles, a single tRNA Lys UUU reads both codons. In mammals, two isoacceptors, tRNA Lys1,2 CUU , decode the AAG codon, and a third, tRNA Lys UUU , translates both codons. The wobble xm 5 s 2 U modification is required for the efficient and specific recognition of both lysine codons, and for translocation on the ribosome. 129 The xm 5 s 2 U 34 modifications, such as bacterial tRNA modification mnm 5 s 2 U 34 , can undergo keto-enol tautomerism ( Fig. 7C ), which allows recognition of both AAA and AAG by expanding the hydrogen bonding capabilities of the nucleoside. Similarly, expanded codon recognition could be achieved via ionization of the nucleoside. In the case of mnm 5 s 2 U, the secondary amine in the sidechain could be positively charged and the thio-group negatively charged. This zwitterion would have the same hydrogen bonding qualities as the tautomer ( Fig. 7C ). 130
Group (VI) elements at the 2 position of U – the influence of atom size
When U is found at tRNA's position 34 it is nearly always modified. Modifications of U at the wobble position include 2-thiouridine (s 2 U) and 2-selenouridine (se 2 U) ( Fig. 8 ) 131 that affect both ribose sugar pucker as well as the glycosidic χ angle resulting in a restrictive reading of codons. Both sulfur as well as selenium have similar chemical properties to the carbonyl oxygen at the C2 position of U. However, the larger size of sulfur and selenium elicit different steric properties on their environment, being approximately 40% and 100% larger than oxygen, respectively ( Fig. 8 ). Also the electronegativity of the substituent atoms (sulfur and selenium) is smaller than that of oxygen. As would be expected, the modified U exhibits a dominant anti conformation. The larger atomic radii of these modifications also affect the nucleoside's sugar pucker particularly when the glycosidic bond angle χ is in the anti conformation. While the ribose of unmodified uridines populate equally the C3′ and C2′ endo conformations, the ribose sugar pucker of s 2 U and se 2 U exist preferentially in the C3′ endo conformation only occasionally switching to the C2′ endo pucker. The large size difference in the Van der Waal radius of the group IV elements bonded to C2 of U forces the sugar to adopt the C’3 endo conformation; a conformation that has been principally associated with increased stability through enhanced stacking. 28 In the case of the unmodified uridine, the carbonyl oxygen at C2 can form a hydrogen bond with the 2′OH group on the ribose thus stabilizing the C2′ endo conformation of the ribose ( Fig. 8A ). The less electronegative S or Se have a reduced propensity for such hydrogen bond interactions, and combined with the steric effects introduced by the larger size of the atoms, these modifications push the sugar pucker distribution toward C3′ endo ( Fig. 8B and 8C ). Thus, the conformation of uridines at wobble position 34 and modified with s 2 and se 2 is anti , C3′ endo , and the base pairing is preferentially to A, rather than wobbling to G.

Sulfur and selenium atomic radii affect sugar pucker when modifying the C2 position of uridine. A. Uridine, B. 2-thiouridine and C. 2-selenouridine nucleosides are shown. All 3 nucleosides are depicted in their anti conformation. The plane defined by atoms C1′-O4′-C4′ of the sugar separates the endo face (the side in which C5′ projects from the plane) from the exo face. In the case of uridine, the sugar pucker is C2′ endo as indicated by atom C2′ in the endo face (above the C1′-O4′-C4′ plane), while in 2-thiouridine and 2-selenouridine, the relative increase in atomic radius of substituent atom X at position C2 and the weakening of the hydrogen bonding interaction between 2′OH and X (where X – O2, S 2 or Se 2 ) shifts the equilibrium toward a C3′ endo conformation.
We and others have extolled the virtues of modifications at wobble position 34 and the invariant purine 3′-adjacent to the anticodon at position 37. Wobble base pairing does not appear to occur at positions other than tRNA position 34 with rare exception such as arthropod mitochondrial translation of the codon AGG as lysine instead of serine (invertebrates) or arginine (standard Universal Genetic Code) in which there is position 35 wobble base pairing, U 35 ○G2. 132 Modifications at tRNA's nucleosides 34 and 37 frame the anticodon and pre-structure the ASL for decoding. In doing so, they reduce energy barriers to conformational change required for ribosomal A-site binding, maintain the translational reading frame and either expand or restrict cognate and wobble codon recognition. In S cerevisiae , loss of either the xm 5 or s 2 modifications of mcm 5 s 2 U 34 increases observation of +1 translational frameshifts, 133 probably caused by a decrease in the affinity of the tRNA for the ribosomal A-site and an impaired translocation. 9 , 129 The nearly ubiquitous t 6 A 37 modification at position 37 in tRNAs responding to codons beginning with A, ANN, enhances base stacking of U 36 with A1 of the codon, as well as prevents intraloop base pairing within the ASL. 115 Thereby, t 6 A 37 facilitates the U-turn and presentation of the anticodon to the codon on the ribosome. t 6 A 37 also facilitates ribosomal codon binding and maintains the translational frame. 26 , 126 Other modifications at position 37, particularly in tRNAs responding to codons beginning with U, UNN, such as wyosine, contribute similarly to base stacking, maintaining the open loop structure and by doing so maintain the translational reading frame. 26 , 126 The fact that the ASL can be stabilized by various purine 37 modifications implies convergent evolutionary pathways that relate not only to the identity of the codon, but also to that of the nucleoside at position 34 and perhaps additionally either 32 or 38/39.
Yet, there appears to be a need for some tRNAs to further modify the anticodon loop for acceptance on the ribosome and codon recognition. For instance, the nucleoside N3-methylcytidine (m 3 C) was first found in tRNA over 50 years ago 134 and appears to be located exclusively at position 32 of the anticodon loop in specific eukaryotic tRNAs. 20 In addition, 2-thiocytidine is found at position 32 of specific prokaryotic tRNAs. The s 2 C 32 modification is present in E. coli and Salmonella enterica tRNA Arg ICG which decodes CGU/C and CGA in the absence of s 2 C 32 , tRNA Arg CCG that decodes CGG, tRNA Arg with the modified anticodon mnm 5 UCU that decodes AGA/G, and tRNA Ser GCU decoding AGC/U. 19 , 39 The presence of s 2 C 32 negates I 34 wobbling to A3. 39 It may affect translational efficiency of rare codons that are intrinsically inefficient in decoding. 19 , 135 The s 2 C 32 modification has also been found in archaeal tRNA. 136 Pseudouridine, Ψ, although present at the anticodon wobble position 34, is another modification found predominantly at positions 32, 38 and 39. 137 Pseudouridine in the anticodon loop apparently increases thermal stability but does not affect ribosome mediated codon binding, nor is its N1 position used for hydrogen bonding. 127
Multiple modifications within the ASL may provide a redundancy to the structure and conformational dynamics not otherwise achievable by 40+ individual tRNAs meeting ribosome acceptance and unique codon recognition. 43 Modification of tRNAs anticodon loop positions 32, 38 or 39, along with positions 34 and 37, in effect complete a triangle of modified nucleosides ( Fig. 9 ). It is interesting that the three point presentation of ASL modifications has modified nucleosides spaced almost equally around the anticodon loop at approximately every other position 32, 34, and 37, or 34, 37 and 39. Looking back at tRNA Gly NCC , the strong anticodon-codon, C•G interactions require no modifications, whereas tRNA Lys UUU , tRNA Tyr NUA , and tRNA Ile NAU have at least three functionally and/or structurally essential modified nucleosides in the ASL. Weak interactions with the codon, such as multiple U•A base pairs, may require far more extensive modifications, where chemically diverse modifications affect similar biologic properties, often through structure.

ASL modification triangle. A combination of 3 positions in the ASL (nucleotides 32 through 38) of tRNAs are commonly modified to expand or restrict codon recognition. Position 32, 34 and 37 (light blue) or position 34, 37 and 38 (dark blue) form vertices of the triangle in which modifications work in a co-surgical manner to achieve desired ribosomal binding affinity of the tRNA to different codons. The anticodon is represented in red and the nucleosides in the ASL in black.
The ribosome bound structures of the tRNA ASLs containing post-transcriptional modifications offer molecular insights into their role in modulating structure, stability and interactions of the tRNA. While the uridine at wobble position 34 is the most heavily modified, positions 32, 37 and 38 also show significant levels of modifications. We have gathered the structures of five tRNAs bound to the ribosome at the A-site, and highlighted the interactions of their modifications ( Fig. 10 ). It is interesting to note that U 34 is primarily modified at C5 adjacent to the base-pairing face and hence the modifications do not interfere with its hydrogen bonding to the codon except in cases where the modifications introduce tautomerism. However, the modifications are either polar (as is true in case of tRNA Leu 138 and tRNA Val 11 ) or charged, (tRNA Lys ), 44 and therefore participate in hydrogen bonding with neighboring bases or charge-charge interactions with the backbone phosphate group. Modifications on the position 37 purine nucleobase are primarily involved in stacking with the base at position 36 and the base to which it is paired in codon recognition on the ribosome. These modifications are either methylations (tRNA Val ) or groups with complex chemistries, like t 6 A 37 , which significantly increases its stacking propensity. Enhanced base stacking of position 37 modified purines provides additional stability to the U-turn structure, and increases the binding strength of the ASL when U 34 pairs with a near-cognate or non-cognate codon.

Ribosome-bound structures of the ASLs of 5 tRNA species with modifications. tRNA modified nucleosides at positions 32, 34 and 37 are influential in creating the architecture of the anticodon domain accepted into the ribosome's A-site for cognate and wobble codon binding. The codon on the mRNA is shown in green, the ASL in cyan with the modifications labeled and the rRNA interactions shown in magenta. Possible interactions of the modified groups are represented using a dashed line. A. In ASL Val UAC bound to codon GUU, (PDB ID: 2UUB), the carboxyl oxygen of cmo 5 U 34 is within hydrogen bonding distance of the amine group on the neighboring A 35 . B. In ASL Leu UAA bound to codon UUG (PDB ID: 2VQF), two of the three oxygens bonded to the sulfur atom in the τm 5 U 34 can form hydrogen bonding interactions with A 35 and A 36 . C. In ASL Lys UUU bound to codon AAA (PDB ID: 1XMQ), there are a salt-bridge between the 5-methylaminomethyl group on U 34 and its phosphate group, an enhanced stacking interaction provided by t 6 A 37 , and a possible hydrogen bond between the threonyl group of t 6 A 37 and A1 on the mRNA. D. ASL Arg2 ICG bound to the codon CGC. The ASL has the two modifications I 34 , m 2 A 37 . E. ASL Arg1 ICG bound to the codon CGC. The ASL has the modifications s 2 C 32 , I 34 . When either of the modifications m 2 A 37 or s 2 C 32 are present, then the tRNA is unable to recognize the rare CGA codon. 39
The cross-loop interaction between the nucleosides at position 32 and 38 at the beginning of the ASL loop ( Fig. 4B ) is characterized by a single or bifurcated hydrogen bond. 139 The strength of this interaction appears to be carefully modulated. For instance, pseudouridine at position 32 uses a water mediated base-backbone interaction to stabilize the interaction between nucleosides 32 and 38. 138 While a stronger interaction, like that of a canonical base pair at this position could lead to a loss in flexibility of the loop domain, a weaker interaction could result in loss of stacking in the ASL resulting in a disruption of the functional U-turn conformation of the ASL. Modifications at the 32 nd and 38 th positions of the ASL may be important as a possible means of modulating the interaction between the two nucleosides and base stacking in the loop.
We hypothesize that the combined chemistries and conformational dynamics of modified nucleosides located at three positions, positions 34, 37 and one other within the ASL loop ( Fig. 9 ) transform the loop architecture and dynamics to that consistent with the constraints that the ribosome places on all tRNAs. It is important that a stable, yet adaptable, U-turn is maintained for presentation of the anticodon resulting in accurate and efficient codon recognition. Multiple modifications mold the anticodon loop architecture of specific tRNAs into thermally stable, malleable triangles of strength recognizable by mRNA-programmed ribosomes ( Fig. 9 ). In order for the architecture of the anticodon's loop to change, an edge of the triangle must collapse, as in an alteration of the 5′-side and U-turn. This is apparent in the three different anticodon loop conformers of tRNA Arg1,2 that are recognized by argininyl-tRNA synthetase with a disrupted U-turn, a solution structure that does not exhibit a U-turn and a conformation on the ribosome in the A-site that has the canonical U-turn. 39
Conjecture about the evolution of site- and chemically specific modifications within tRNA's ASL domain relative to their functions can yield insights into their future, investigator-designed applications. RNA polymerases, with few exceptions, in vivo and in vitro , do not accept modified nucleoside triphosphates as substrates for transcription, and if they did the modification would be randomly placed throughout the transcript in response to the template. Therefore, it is feasible to hypothesize that the evolution of post-transcriptionally modified nucleosides occurred after the appearance of proteins that could catalyze their limited existence at specific sites to which each modification would contribute chemistry and conformation to that tRNA's function in translation. In acknowledging today's understanding of the manner in which anticodon domain modifications both expand and restrict recognition of cognate and wobble codons, but not near-cognate codons, it is difficult to comprehend life in which new amino acids would be introduced with limited numbers of tRNAs and necessitating 2-fold degenerate codons. For instance, without the restrictions imposed by modifications, one could conceive of asparagine and lysine, aspartic and glutamic acids, and perhaps arginine and serine being mistakenly incorporated. More likely, tRNAs with unmodified anticodon domains much like the very limited number of 22 tRNAs transcribed in mammalian mitochondria decoded 4-fold degenerate codon boxes without error, but perhaps at reduced efficiencies. In contrast, cytoplasmic and mitochondrial translation of codons from 2-fold degenerate codon boxes ( Fig. 1 ) preceded in the absence of modification with significantly reduced translational fidelity and lack of efficiency until restrictive modifications had evolved. If tRNA anticodon domain modifications facilitated the accurate and efficient entry of new amino acids through split codon boxes in the evolution of proteins as we know them, then why couldn't the emergence of still other amino acid incorporations continue by re-appropriating sense codons? 140 , 141 The use of 4-fold degenerate sense codons to incorporate new amino acids would require the splitting of 4-fold degenerate codons. Perhaps, this could be accomplished by removal of cytoplasmic tRNAs from redundant codon recognition (which does not occur in the mitochondria), restructuring the modification enzyme recognition of specific tRNA species and aminoacyl-tRNA synthetase recognition of tRNA and the non-natural amino acid substrates. The introduction of novel amino acids into proteins portends applications to biomaterial science and medicine. New investigator-designed, protein-based materials and potential therapeutics would become available through biomanufacturing.
In conclusion, The Wobble Hypothesis is being revised continually for as we learn more every year about modified nucleoside chemistry, structure and function and how it reflects on RNA chemistry, structure and function, we understand the subtleties of translation being able to manipulate and apply them. We have discussed the different instances in which wobble base pairs are used to expand/restrict codon recognition by tRNA's anticodon with modified and unmodified wobble position 34, and the importance of modified nucleoside positions outside of anticodon, 32, 37 and 38 ( Fig. 11 ). Wobble and non-canonical base pairs in the ASL domain of tRNA are integral to the translation of the genetic code. They are modulated either by unique structural and dynamic features introduced in a sequence dependent manner by unmodified bases (e.g., tRNA Gly with C 32 /U 32 ) or by nucleoside modifications that can expand or restrict decoding capacity of tRNAs by stabilizing or disrupting native interactions at the decoding center of the ribosome. An in-depth understanding of the origins and mechanisms of the variety of ways by which wobbling is achieved will equip us with the tools to tap into this often overlooked potential of using modified nucleoside contributions to translation for therapeutics and other applications.

Wobble map. Anticodon domain modified nucleosides play an important role in the wobbling of tRNA's codon recognition, some expanding codon recognition to synonymous codons while others restrict recognition. The different cases of wobbling in the presence and absence of modifications as discussed in this paper, are shown. The black arrow represents no modifications, whereas a red arrow indicates the presence of one or more modifications. The tRNAs are shown in blue, the anticodons are indicated on the arrow and codons decoded by the tRNA bearing the anticodon are enclosed in white boxes.
Disclosure of potential conflicts of interest
The authors report no conflict of interest.
Research conducted by the authors is supported by grants to P.F.A. from the Department of Defense Medical Research Program under grant number W81XWH-16-1-0428, PR151248, the National Science Foundation under grant number CHE1407042 and the National Institutes of Health, National Institute of General Medical Sciences under grant number 5R01GM110588-03 to Manal Swairjo, P.F.A. co-investigator.

What is the wobble theory?
Wobble theory: the crick wobble hypothesis attributes the phenomenon of codon degeneracy to an imprecise pairing of the codon's third base and the anticodon's first base. the pairing features of several wobble bases, both seen and undiscovered, are investigated in this theoretical work. the genetic code is degenerate, which means that one amino acid is coded by several codons. to explain the degeneracy of codons in the third position of the codon. a wobble is defined as an unstable movement or a quiver in the voice..

What is the wobble effect?

Explain 'Wobble hypothesis' with the help of a suitable diagram.
Wobble hypothesis states the degeneracy of the genetic code. the pairing of the third base varies according to the base at the third position, for example, g may pair with u. the conventional pairing (a = u, g = c) is called watson-crick pairing and the second abnormal pairing is called wobble pairing..

Biology Notes Online
Iyo Wobble Hypothesis - Tsanangudzo, Tsananguro, Kukosha
Zviri Mukati
Chii chinonzi Wobble Hypothesis?
Iyo Wobble Hypothesis, yakakurudzirwa naFrancis Crick muna 1966, inopa tsananguro yekuderera kweiyo genetic code. Degeneracy inoreva chokwadi chekuti macodon akawanda anogona kukodha kune imwechete amino acid. Zvinoenderana neWobble Hypothesis, kubatanidza chaiko pakati pemabhesi ekodon uye anticodon yetRNA inoitika chete kune maviri ekutanga mabhesi ekodon. Nekudaro, iyo pairing pakati pechitatu base yekodon uye anticodon inogona kuratidza kumwe kuchinjika kana "kuzununguka."
Mune mamwe mazwi, nheyo yechitatu yekodon uye anticodon inogona kuzununguka kana kufamba isina kugadzikana, ichibvumira kune isiri-standard base pairing. Ichi chiitiko chinogonesa imwechete tRNA molecule kuziva uye kusunga kune akawanda macodon, zvisinei nekusiyana muhwaro hwechitatu. Nekuda kweizvozvo, kunyangwe paine macodon makumi matanhatu nerimwe anoisa maamino acids, huwandu hwetRNA mamorekuru hwakadzikira zvakanyanya (kutenderera makumi mana) nekuda kwekuzununguka.
Wobble base pairs vanoita basa rakakosha muRNA yechipiri chimiro uye yakakosha pakududzira kwakaringana kweiyo genetic code. Iwo mana mahombe akadzikama base pairs anoti guanine-uracil (GU), hypoxanthine-uracil (IU), hypoxanthine-adenine (IA), uye hypoxanthine-cytosine (IC). Aya mawobble base pairs anoratidza kugadzikana kwe thermodynamic kuenzaniswa neiyo yeWatson-Crick base pairs.
Mune genetic code, kune makumi matanhatu nemana anokwanisika macodons, kubva mairi matatu ari stop codons anogumisa kushandura. Kana canonical Watson-Crick base pairing yaidiwa kune yega yega codon, zvaizoda 64 akasiyana marudzi emamorekuru etRNA. Zvisinei, zvipenyu zvizhinji zvine marudzi asingasviki makumi mana neshanu etRNA. Iyo Wobble Hypothesis inotsanangura kuti mamwe mamorekuru etRNA anogona sei kuziva akawanda anofanana macodon, ayo encode yakafanana amino acid.
Crick akakurudzira kuti iyo 5 ′ base pane anticodon, iyo inosunga kune iyo 3 ′ base pane mRNA codon, inonyanya kuchinjika munzvimbo kana ichienzaniswa nemamwe mabhesi maviri. Uku kuchinjika kunobvumira kusiri-standard base pairing uye diki conformational gadziriso, zvichikonzera iyo yakazara pairing geometry yetRNA anticodons.
Pakazere, iyo Wobble Hypothesis inopa nzira yekuzvidavirira yekuderera kweiyo genetic code uye kugona kwenhamba shoma yemamorekuru etRNA kuziva uye kusunga kune akawanda macodon. Kuzunguzika kwechigadziko chechitatu kunobvumira kushanduka kukuru kwemajini coding uku uchichengetedza huchokwadi hwekushandura.
Tsanangudzo yeWobble Hypothesis
Iyo Wobble Hypothesis inokurudzira kuti hwaro hwechitatu hwekodon uye anticodon ye tRNA inogona kuratidza kuchinjika kana "kuzununguka" munheyo yavo pairing, zvichibvumira imwechete tRNA molecule kuziva uye kusunga kune akawanda codon, zvichikonzera kuderera kweiyo genetic code.
Iyo Wobble Hypothesis
Iyo Wobble Hypothesis inokurudzira kuti hwaro pa5 ′ kumagumo kweanticodon muTRNA haina kumanikidzwa munzvimbo semamwe mabhesi maviri. Uku kuchinjika kunobvumira kuti igadzire ma hydrogen zvisungo ane akawanda mabhesi ari pa 3 ′ kumagumo ekodon mumRNA. The wobble hypothesis inoti:
- Mabhesi maviri ekutanga ekodon uye mabhesi anoenderana mune anticodon fomu yakajairika Watson-Crick base pairs.
- Nzvimbo yechitatu mucodon inotevera mitemo isinganyanyi kuomesera, inotungamira kune isiri-canonical pairing kana "kuzununguka."
- Iyo yakazorora base-pairing inodiwa pachinzvimbo chechitatu inogonesa imwechete tRNA molecule kuti ibatane neinopfuura imwe mRNA katatu.
- Mitemo yakananga yekupeta pairing ndeiyi: U ari pachinzvimbo chekutanga cheanticodon anogona kuziva A kana G mucodon, G anogona kuziva U kana C, uye ini (inosine) ndinogona kuziva U, C, kana A.
- Aya maitiro akatungamira Francis Crick kuti ataure iyo wobble hypothesis, iyo inotsanangura inochinjika base-pairing kudyidzana kunoonekwa muiyo genetic code.
Iyo coding chaiyo yeiyo genetic code inonyanya kutsamira pamabhesi maviri ekutanga ekodon, ayo anoumba yakasimba Watson-Crick base pairs ne anticodon ye tRNA. Nucleotide yekutanga mu anticodon inosarudza kuti ingani nucleotides iyo tRNA inogona kusiyanisa pakuverenga codon mu 5' kusvika 3 'kutungamirira.
Kana iyo nucleotide yekutanga muanticodon iri C kana A, kubatanidza kwacho kwakananga, uye chete codon chaiyo inogona kuzivikanwa neiyo tRNA. Nekudaro, kana iyo nucleotide yekutanga iri U kana G, iyo pairing haina kujeka, ichibvumira kuchinjika pakati pezvigadziko zviviri mukodon. Panyaya yeinosine seyokutanga nucleotide, inoratidza huroyi hwechokwadi, huchiita kuti ibatane nechero yezvigadziko zvitatu mukodonyo yekutanga.
Nekuda kwekujeka kwemanyucleotide maviri ekutanga mucodon, kana amino acid yakarongedzerwa nemaanticodon akawanda uye iwo maanticodon akasiyana munzvimbo yechipiri kana yechitatu (yekutanga kana yechipiri chinzvimbo mucodon), tRNA yakasiyana inodiwa kune imwe neimwe. anticodon.
Kuti ugutse macodon ese anobvira (61 kusanganisa matatu anomira macodon), mashoma makumi matatu nemaviri tRNA mamorekuru anodiwa. Izvi zvinosanganisira 32 tRNAs yeamino acids uye imwe yekutanga codon.
Iyo Wobble Hypothesis inopa kunzwisiswa kwakakosha kwekushanduka uye kuderera kweiyo genetic code, ichibvumira nhamba shoma yemamorekuru etRNA kuti azive uye asunge kune akawanda macodon apo achichengetedza huchokwadi hweprotein dudziro.

Wobble base pairs

Wobble base pairs ndiwo chaiwo pairings pakati pemanucleotides muRNA mamorekuru anotsauka kubva kune yakajairwa Watson-Crick base pair mitemo. Ivo vanoita basa rakakosha muRNA chimiro uye dudziro. Heano mamwe mapoinzi akakosha nezve wobble base pairs:
- Wobble base pairs inosanganisira isiri-yakajairwa pairings pakati pemanucleotides munzvimbo chaidzo.
- Iwo mana mahombe epasi peya akaonekwa muRNA anoti guanine-uracil (GU), hypoxanthine-uracil (IU), hypoxanthine-adenine (IA), uye hypoxanthine-cytosine (IC).
- Kushandiswa kwe "I" ye hypoxanthine inochengetedza kuwirirana mu nucleic acid nomenclature sezvo hypoxanthine ndiyo nucleobase ye inosine.
- Pakati pewobble base pairs, inosine inoratidza hunhu hwakakosha. Kana inosine iripo seyokutanga nucleotide muanticodon ye tRNA, inogona kubatana nechero yezvigadziko zvitatu (adenine, cytosine, kana uracil) mukodononi inoenderana nemRNA.
- Kugona kweInosine kuzununguka kunobvumira molecule imwe chete yetRNA ine inosine-ine anticodon kuti ione macodon akawanda, kuwedzera kuchinjika kweiyo genetic code.
Wobble base pairs inosuma mwero wekusiyana-siyana mukudyidzana kweRNA, kunyanya panguva yekushandura. Vanogonesa nhamba yakaderedzwa yemamorekuru etRNA kuti azive akawanda macodon, achibhadhara kuderera kweiyo genetic code. Iyo wobble hypothesis, iyo inotsanangura izvi zvisiri-yakajairwa pairings, inopa nzwisiso mukubudirira uye kurongeka kweprotein synthesis.

Tsanangudzo Pfupi yeThe Wobble Hypothesis
Ngatinyure muWobble Hypothesis nhanho nhanho.
Iyo Wobble Hypothesis ipfungwa inotsanangura kuti iyo genetic code, yakachengetwa muchimiro che nucleotides, inoshandurwa kuita mapuroteni ne ribosome. Kuti tinzwisise pfungwa iyi, tinoda kuziva zvishoma nezvemakodon uye anticodons.
Macodon anotevedzana matatu nucleotides mumRNA, uye imwe neimwe codon inoenderana neyakananga amino acid kana chiratidzo chekumira. Kune rimwe divi, maanticodon anotevedzana matatu nucleotides mutRNA anosunga kune codon panguva yeprotein synthesis.
Zvinoenderana neWobble Hypothesis, hwaro pa5 ′ kumagumo kweanticodon haina kuganhurirwa mukubatanidza kwayo semamwe mabhesi maviri. Izvi zvinoreva kuti mabhesi maviri ekutanga ekodon uye anticodon anoumba zvakajairwa hydrogen bond pairs, achitevera yakajairika base-pairing mitemo (A neU, G neC). Zvisinei, panzvimbo yechitatu yekodon, mitemo yakasununguka, uye kwete-canonical pairing inogona kuitika.
Mune mamwe mazwi, hwaro hwechitatu hwekodon uye hwaro hwekutanga hweanticodon hunogona kuumba "kuzununguka" pairs dzisinganyatso kutevedzera AU neGC base-pairing mitemo. Uku kuchinjika kunobvumira anticodon yeimwe tRNA molecule kuziva uye kusunga kune inodarika codon imwe ine nucleotide sequences dzakasiyana panzvimbo yechitatu.
Kuti ndikupe iwe mimwe mienzaniso yemitemo ye wobble pairing:
- Kana hwaro hwekutanga hweanticodon iri U, inokwanisa kuziva macodon ane A kana G sehwaro hwechitatu.
- Kana hwaro hwekutanga hweanticodon iri G, inogona kuziva macodon ane U kana C sehwaro hwechitatu.
- Kana hwaro hwekutanga hweanticodon ndini (Inosine), inogona kuziva macodon ane U, C, kana A sehwaro hwechitatu.
Nekubvumidza iyi "kuzununguka" kana kudzikama base-pairing, sero rinogona kudzikisa huwandu hwetRNA mamorekuru anodiwa paprotein synthesis. Inopa kushanduka uye kushanda zvakanaka mukushandura genetic code.
Saka, muchidimbu, iyo Wobble Hypothesis inokurudzira kuti nheyo yechitatu yekodon uye nheyo yekutanga ye anticodon inogona kuumba isiri-standard base pairs, zvichiita kuti iwedzere kushanduka yemitemo ye-base-pairing panzvimbo yechitatu yekodon. Uku kuchinjika kunobvumira imwechete tRNA molecule kuziva uye kusunga kune akawanda macodon ane akasiyana nucleotide sequences panzvimbo yechitatu, optimizing protein synthesis.
Kukosha kweWobble Hypothesis
Iyo wobble hypothesis inobata kukosha kwakakosha mukunzwisisa kugona uye kurongeka kweprotein synthesis. Heano mapoinzi akakosha anoratidza kukosha kweiyo wobble hypothesis:
- Yakakura chaiyo ine mashoma tRNAs: Miviri yedu ine nhamba shoma yemamorekuru etRNA. Iyo wobble hypothesis inobvumira tRNA imwe chete kuziva macodon akawanda nekuda kweasina-standard pairings panzvimbo yekuzununguka. Uhu hwakapamhama hunogonesa kuturikira kwakanaka netRNAs diki.
- Kufambisa mabasa ebhayoloji: Wobble base pairs have been extensively studied in organisms such as Escherichia coli ( E. coli ), demonstrating their role in various biological processes. They contribute to the accuracy of translation and protein synthesis.
- Kuenzanisa thermodynamic kugadzikana: Wobble base pairs inoratidza thermodynamic kugadzikana yakafanana neWatson-Crick base pairs. Uku kugadzikana kunovimbisa kutendeseka kweRNA yechipiri zvimiro uye inosimudzira yakavimbika dudziro yeiyo genetic code.
- Yakakosha kune RNA yechipiri chimiro: Wobble base pairs vanoita basa rakakosha mukuumbwa kweRNA yechipiri zvimiro. Ivo vanobatsira mukugadzikana uye kupeta kwemamorekuru eRNA, zvichiita kuti kushanda zvakanaka kweRNA mumaserura akasiyana siyana.
- Kukurumidza kuparadzaniswa uye mapuroteni synthesis: Iyo wobble base pairing inobvumira kukurumidza kuparadzaniswa kwetRNA kubva mRNA panguva yekushandura. Uku kukurumidza kuparadzaniswa kunokurudzira kushanda kweprotein synthesis nekufambisa kufamba kweribosomes pamwe nemRNA template.
- Kuderedza zvikanganiso mukududzira kodhi kodhi: Kuvapo kwekuzununguka kunoderedza kukanganisa kwezvimwe zvikanganiso mune genetic code. Kana codon ikasaverengwa panguva yekunyorwa, kuzununguka kunobvumira tRNA kuti irambe ichiziva uye kushandura nemazvo kodon, ichichengetedza kutevedzana kweamino acid panguva yeprotein synthesis. Izvi zvinoderedza kukanganisa kunogona kuitika kubva pane dzimwe nguva kukanganisa mukuverenga genetic code.
Mibvunzo paWobble Hypothesis
Chii chinotsanangurwa neWobble Hypothesis? a) Chimiro cheDNA b) Kudzokororwa kweDNA c) Iko kuchinjika mukubatanidza kwechitatu hwaro hwekodon d) Kuumbwa kwemapuroteni
Ndiani akakurudzira Wobble Hypothesis? a) Rosalind Franklin b) James Watson c) Francis Crick d) Maurice Wilkins
Zvinoenderana neWobble Hypothesis, ndeipi chinzvimbo chekodon chinoratidza kuchinjika mu base pairing? a) Kutanga b) Chechipiri c) Chechitatu d) Chechina
Ndechipi chigadziko chinogona kubatana nemabhesi akawanda maererano neWobble Hypothesis? a) Adenine b) Cytosine c) Guanine d) Thymine
Iyo Wobble Hypothesis inobatsira kutsanangura chikonzero nei: a) Kune akawanda macodon kupfuura amino acids b) Kune akawanda amino acids kupfuura makodon c) Makodoni anogara akareba d) DNA ine tambo mbiri
Ndechipi chezvinotevera CHISI chibvumirano chinozunguzika? a) GU b) AU c) IA d) CG
Iyo Wobble Hypothesis inoderedza kudiwa kwe: a) Multiple DNA tambo b) Mhando dzakawanda dzeamino acids c) Mhando dzakawanda dzetRNAs d) Mhando dzakawanda dze ribosomes
Inosine (I) inogona kubatanidza neipi yezvigadziko zvinotevera? a) Adenine b) Cytosine c) Zvose Adenine uye Cytosine d) Kwete Adenine kana Cytosine
Iyo Wobble Hypothesis inonyanya kubatanidzwa ne: a) DNA replication b) Chinyorwa c) Translation d) DNA kugadzirisa
Iko kuchinjika mu base pairing, sezvakatsanangurwa neWobble Hypothesis, inoitika pakati: a) mRNA codon uye DNA template b) mRNA codon uye tRNA anticodon c) tRNA anticodon uye DNA template d) tRNA anticodon uye ribosomal RNA
Chii chinonzi wobble hypothesis?
Iyo wobble hypothesis inokurudzira kuti hwaro pa5 ′ kumagumo eanticodon mu tRNA haina kunyatso kubatanidzwa neiyo inoenderana hwaro mumRNA codon, ichibvumira isiri-standard kana wobble base pairings.
Sei ichinzi "wobble" hypothesis?
Inonzi "wobble" hypothesis nekuti hwaro panzvimbo yekuzununguka haina kuvharirwa munzvimbo semamwe mabhesi maviri ari muanticodon, ichibvumira kuzununguka kana kufamba zvisina kugadzikana uye kugadzira asiri-standard base pairs.
Ko iyo wobble hypothesis inotsanangura sei kuderera muiyo genetic code?
Iyo wobble hypothesis inoratidza kuti yakasununguka base-pairi yemitemo panzvimbo yechitatu yekodon inobvumira imwechete tRNA molecule kuziva inodarika codon imwe. Izvi zvinokonzeresa kuderera kana kudzokororwa kweiyo genetic code.
Ndeapi makuru wobble base pairs?
Iwo mahombe obble base pairs anoti guanine-uracil (GU), hypoxanthine-uracil (IU), hypoxanthine-adenine (IA), uye hypoxanthine-cytosine (IC).
Sei hypoxanthine ichishandiswa mu wobble base pairs?
Hypoxanthine inoshandiswa kumiririra wobble base pairs nekuti ndiyo nucleobase yeinosine, iyo inoratidza hunhu chaihwo hwekuzungaira nekubvumira kubatanidza nemabhesi akawanda mukodononi yekutanga.
Ko iyo wobble hypothesis inobata sei protein synthesis?
Iyo wobble base pairing inobvumira kukurumidza kuparadzaniswa kwetRNA kubva mRNA panguva yeprotein synthesis, kufambisa kufamba kwema ribosomes uye kuwedzera kugona kwekushandura.
Ndeipi basa rinoitwa wobble base pairs muRNA yechipiri chimiro?
Wobble base pairs akakosha muRNA yechipiri chimiro. Ivo vanobatsira mukugadzikana uye kupeta kwemamorekuru eRNA, zvichipesvedzera chimiro chavo chose uye basa.
Ko iyo wobble hypothesis inokanganisa kurongeka kweiyo genetic code?
Hongu, iyo wobble hypothesis inobatsira kuderedza zvikanganiso mukududzirwa kweiyo genetic code. Inovimbisa kuti kunyangwe paine kusawirirana panzvimbo yekuzununguka, iyo chaiyo amino acid inogona kuramba ichibatanidzwa panguva yeprotein synthesis.
Ko iyo wobble hypothesis inobata sei huwandu hwemamorekuru etRNA anodiwa?
Iyo wobble hypothesis inobvumira imwechete tRNA molecule kuti ione akawanda macodon nekuda kweasina-standard pairings. Izvi zvakapamhama zvinodzikisira huwandu hweakasarudzika tRNA mamorekuru anodiwa pakushandura.
Ko fungidziro yekuzununguka yakabatsira sei pakunzwisisa kwedu mamolecular biology?
Iyo wobble hypothesis yakapa nzwisiso mukubudirira uye kurongeka kweprotein synthesis, RNA chimiro, uye kushanda kweiyo genetic code. Zvakawedzera kunzwisisa kwedu kuti maseru anokwirisa sei zviwanikwa uye kuchengetedza kuvimbika muhurongwa hwakaoma hwekushandura ruzivo rwemajini kuita mapuroteni anoshanda.
- https://microbenotes.com/the-wobble-hypothesis/
- https://en.wikipedia.org/wiki/Wobble_base_pair
- https://teaching.ncl.ac.uk/bms/wiki/index.php/Wobble_Hypothesis
- https://link.springer.com/referenceworkentry/10.1007%2F978-3-642-11274-4_1692
- https://www.biologydiscussion.com/genetics/wobble-hypothesis-with-diagram-genetics/65163
Related Posts
- Chii chinonzi Dihybrid Cross? Mienzaniso, Matanho, Kukosha
- Mitemo yeChargaff - Yekutanga uye Yechipiri Mutemo
- Karyotype uye Idiogram - Tsanangudzo, Maitiro, Matanho, Zvishandiso
- Codon Chati uye Codon Tafura
- Deletion Mutation – Tsanangudzo, Zvikonzero, Mechanism, Mienzaniso
Leave a Comment kukanzura mhinduro
Sevha zita rangu, email, uye webhusaiti mune ino browser kwenguva inotevera ini pandinopindura.
Nzvimbo iyi inoshandisa Akismet kuderedza spam. Dzidza kuti sei deta yako inoshandiswa .

- Online Class
- Ask Doubt on Whatsapp
- Search Doubtnut
Write short note on wobble hypothesis Solution in Odia
More from this Exercise

Step by step video & image solution for Write short note on wobble hypothesis by Biology experts to help you in doubts & scoring excellent marks in Class 12 exams.
Related Playlists
MOLECULAR BASIS OF INHERITANCE
MICROBES IN HUMAN WELFARE
ORGANIC EVOLUTION
Similar Questions
One of the following is true with respect to AUG :
Describe the transiation of prokaryotes.
Organisms called methanogens are most abundant in a
Select the incorrect pair
Wobble hypothesis establishes :
Wobble hypothesis was proposed by :
Wobble hypothesis was given by
Describe the wobble hypothesis.
वोबल परिकल्पना क्या होती है?
Wobble hypothesis _________
वॉबल परिकल्पना का प्रतिपादन किसने किया था
Wobble hypothesis was given by :
Wobble hypothesis explain
Write a short note on degeneracy of genetic code or wobble hypothesis.
Wobble hypothesis was proposed by
ARIHANT PRAKASHAN - MOLECULAR BASIS OF INHERITANCE - TOPIC -2- TOPIC TEST 2
The codon for anticodon 3^'-UUA-5^' is :
Which gene is responsible for the transcription of B-galactosidases?
If a cell is treated with a chemical that blocks nucleic acid synthesi...
Fill In The blank : Aminoacyl synthetase enzyme take part in ………………….
The ......... codes for the repressor in lac operon.
The movement of a ribosome from 5' -3' end of mRNA to recognise all co...
RNA can give rise to DNA through the enzyme ……….
Write a short note on features of human genome project.
Write short note on wobble hypothesis
Degenerate codon and Nonsense codon.
IMAGES
VIDEO
COMMENTS
There are more than one codon for one amino acid.This is called degeneracy of genetic code. To explain the possible cause of degeneracy of codons, in 1966, Francis Crick proposed "the Wobble hypothesis".. According to The Wobble Hypothesis, only the first two bases of the codon have a precise pairing with the bases of the anticodon of tRNA, while the pairing between the third bases of ...
The Wobble Hypothesis, proposed by Francis Crick in 1966, provides an explanation for the degeneracy of the genetic code. Degeneracy refers to the fact that multiple codons can code for the same amino acid. According to the Wobble Hypothesis, the precise pairing between the bases of the codon and the anticodon of tRNA occurs only for the first ...
Here are some examples of the wobble hypothesis in action: Arginine: The amino acid arginine is coded by six different codons: CGU, CGC, CGA, CGG, AGA, and AGG. However, no six different tRNA molecules correspond to each of these codons. Instead, one tRNA molecule with the anticodon 3′-CCU-5′ can recognize the codons CGU, CGC, and CGA ...
In this article we will discuss about the concept of wobble hypothesis. Crick (1966) proposed the 'wobble hypothesis' to explain the degeneracy of the genetic code. Except for tryptophan and methionine, more than one codons direct the synthesis of one amino acid. There are 61 codons that synthesise amino acids, therefore, there must be 61 ...
Wobble Base Pair The Wobble Hypothesis Genetic Code Study Notes: The Wobble Rules Crick also proposed a set of rules that govern the possible wobble base pairs, based on the geometry and hydrogen bonding patterns of the nucleotides involved. The rules are as follows: These rules imply that some codons for the same amino acid […]
Search for: 'wobble hypothesis' in Oxford Reference ». A theory to explain the partial degeneracy of the genetic code due to the fact that some t-RNA molecules can recognize more than one codon. The theory proposes that the first two bases in the codon and anticodon will form complementary pairs in the normal antiparallel fashion.
F.H.C. Crick, in 1965 proposed a hypothesis called Wobble hypothesis to explain this phenomenon. He discovered that if U is present at first position of anticodon, it can pair with either A or G at the third position of codon. Similar is the case with G and I (I = inosine is a modified base in tRNA) found in anticodon. The pairing relationships ...
The wobble hypothesis states that the third position (3') of the codon on mRNA and the first position (5') of the anticodon on tRNA are bound less tightly than the other pair and therefore, offer unusual base combinations. This phenomenon give rise to: The multiple codes for a given amino acid ( degeneracy) Possible suppression of point ...
A code word or codon is the set of nucleotides that specifies one amino acid. By genetic code, one refers to the collection of sequences of bases (codons) that correspond to each amino acid and translation signals. Regarding the possible size of a codon, we can consider George Gamov's (1954) traditional yet rational explanation.
Wobble hypothesis tRNA wobble, Wobble position. A property of the genetic code in which codons that differ in the third position (wobble position) can specify the same tRNA/amino acid.
Wobble base pairing enables the decoding of two or more codons by the same tRNA. Certain modified bases, for example inosine (I) can extend or restrict the degree of flexibility in the range of mRNA:tRNA interactions. Codons that are decoded by wobble base interactions are processed at a slower rate in the ribosome.
A theory to explain the partial degeneracy of the genetic code due to the fact that some t-RNA molecules can
The Wobble Hypothesis •The first two bases of the codon make normal (canonical) H- bond pairs with the 2nd and 3rd bases of the anticodon •At the remaining position, less stringent rules apply and non- canonical pairing may occur •The rules: first base U can recognize A or G, first base G can recognize U or C, and first base I can recognize U, C or A (I
eg. Inosine (I) can pair with A, C, or U. This base is called a Wobble base or fluctuating base. Wobble occurs at position 1 of the anti-codon and position 3 of the codon. The wobble hypothesis states that the third position (3') of the codon on mRNA and the first position (5') of the anti-codon on tRNA are bound less tightly than the other ...
A theory proposed to explain the partial degeneracy of the genetic code in that some t-RNA molecules can recognize more than one ...
Wobble and Superwobble. In most cases, more than one triplet codon can specify an amino acid—at one extreme, leucine can be encoded by any of six nucleotide triplets. Degenerate codons tend to vary at the third position, which was the basis for Francis Crick's wobble hypothesis: Each codon must be recognized by its cognate transfer RNA (tRNA ...
A simple post-transcriptional modification of tRNA, deamination of adenosine to inosine at the first, or wobble, position of the anticodon, inspired Francis Crick's Wobble Hypothesis 50 years ago. Many more naturally-occurring modifications have been elucidated and continue to be discovered. The post-transcriptional modifications of tRNA's ...
Codon-anticodon interaction results in the formation of a short segment of the double helix. In the case of some codons, opposite to uracil in the third position of the codon there is inosine in the anticodon. Crick's wobble hypothesis [1] allows a solution for this pair only by wobbling the third nucleotides of the codon and the anticodon.
An alternative to the wobble hypothesis has been proposed. The facts revealed in this study offer a new insight into physical mechanisms of the functioning of the genetic code. The wobble hypothesis, which was first proposed more than 40 years ago [1], can explain two events: 1) formation of the uracil guanine pair as a result of codon ...
The pairing features of several wobble bases, both seen and undiscovered, are investigated in this theoretical work. The genetic code is degenerate, which means that one amino acid is coded by several codons. To explain the degeneracy of codons in the third position of the codon. A wobble is defined as an unstable movement or a quiver in the voice.
Explain 'Wobble hypothesis' with the help of a suitable diagram. Wobble hypothesis states the degeneracy of the genetic code. The pairing of the third base varies according to the base at the third position, for example, G may pair with U. The conventional pairing (A = U, G = C) is called Watson-Crick pairing and the second abnormal pairing is ...
The Wobble Hypothesis, proposed by Francis Crick in 1966, provides an explanation for the degeneracy of the genetic code. Degeneracy refers to the fact that Zvashandiswa kugutsikana
Watch complete video answer for "Write short note on wobble hypothesis" of Biology Class 12th. Get FREE solutions to all questions from chapter MOLECULAR BASIS OF INHERITANCE.